You might also like
- RL 350 PDFDocument150 pagesRL 350 PDFBhumika89% (9)
- Ready-Mixed White and Yellow Traffic Paints: Standard Specification ForDocument3 pagesReady-Mixed White and Yellow Traffic Paints: Standard Specification ForFRANZ RICHARD SARDINAS MALLCONo ratings yet
- Process Intensification in Lactic Acid ProductionDocument11 pagesProcess Intensification in Lactic Acid ProductionRoxana StoicaNo ratings yet
- Astrological BangleDocument16 pagesAstrological BangleVeeru Kuppast100% (1)
- Experiment 1 (Introduction)Document16 pagesExperiment 1 (Introduction)Msfaeza HanafiNo ratings yet
- Iron Kinetics and Laboratory AssessmentDocument4 pagesIron Kinetics and Laboratory AssessmentJohnree A. EvangelistaNo ratings yet
- Types of Waterproofing Methods For BuildingsDocument6 pagesTypes of Waterproofing Methods For BuildingsKrugerNo ratings yet
- Aromatic CompoundDocument24 pagesAromatic CompoundLegacy MusicNo ratings yet
- Fermentation 04 00045Document12 pagesFermentation 04 00045hmossNo ratings yet
- Acid Lactic Din Xilooligozaharide Cu Rhizopus Oryzae PDFDocument8 pagesAcid Lactic Din Xilooligozaharide Cu Rhizopus Oryzae PDFComan GigiNo ratings yet
- 4.0 Mardawati 2019Document9 pages4.0 Mardawati 2019Bandita DashNo ratings yet
- Evaluation of The Alcoholic Fermentation Kinetics of Enzymatic Hydrolysates From Sugarcane Bagasse (Saccharum Officinarum L.)Document9 pagesEvaluation of The Alcoholic Fermentation Kinetics of Enzymatic Hydrolysates From Sugarcane Bagasse (Saccharum Officinarum L.)Felipe SouzaNo ratings yet
- Biotechnology For Biofuels: Yield-Determining Factors in High-Solids Enzymatic Hydrolysis of LignocelluloseDocument10 pagesBiotechnology For Biofuels: Yield-Determining Factors in High-Solids Enzymatic Hydrolysis of LignocelluloseAlfonsius Dimas AdityoNo ratings yet
- Journal Pone 0107143Document6 pagesJournal Pone 0107143suryasanNo ratings yet
- Optimization of Very High Gravity Fermentacion Process For Ethanol Production From Industrial Sugar Beet SyrupDocument9 pagesOptimization of Very High Gravity Fermentacion Process For Ethanol Production From Industrial Sugar Beet SyrupCAMPOS CASIMIRO ESTELITANo ratings yet
- Factors Affecting Ethanol Fermentation Using SaccharomycesDocument7 pagesFactors Affecting Ethanol Fermentation Using SaccharomycesLl FfNo ratings yet
- A Review On The Current Developments in Continuous Lactic Acid Fermentations and Case Studies Utilising Inexpensive Raw MaterialsDocument10 pagesA Review On The Current Developments in Continuous Lactic Acid Fermentations and Case Studies Utilising Inexpensive Raw MaterialsIntanFauziahNo ratings yet
- Chap1 - 43669Document44 pagesChap1 - 43669Giovani Renato ZontaNo ratings yet
- A Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteDocument11 pagesA Novel Cell Factory For Efficient Production of Ethanol From Dairy WasteVitor ArsenioNo ratings yet
- Biotechnology Progress - 2008 - Lenihan - Developing An Industrial Artemisinic Acid Fermentation Process To Support TheDocument7 pagesBiotechnology Progress - 2008 - Lenihan - Developing An Industrial Artemisinic Acid Fermentation Process To Support The774hzz9zd8No ratings yet
- Rhodosporidium Toruloides Y4 in Fed-Batch Culture: High-Density Cultivation of Oleaginous YeastDocument8 pagesRhodosporidium Toruloides Y4 in Fed-Batch Culture: High-Density Cultivation of Oleaginous YeastAnderlone SidabutarNo ratings yet
- Fermentation UthrDocument12 pagesFermentation UthrJher OcretoNo ratings yet
- Jurnal Allyza Format PertanikaDocument18 pagesJurnal Allyza Format PertanikaPapanya AyuNo ratings yet
- Biomolecules: Lipases Immobilization For Effective Synthesis of Biodiesel Starting From Coffee Waste OilsDocument21 pagesBiomolecules: Lipases Immobilization For Effective Synthesis of Biodiesel Starting From Coffee Waste Oilsjuan diazNo ratings yet
- Chapter 2 - FruitwastesDocument23 pagesChapter 2 - FruitwastesJusztinAquinoNo ratings yet
- Pretreatment and Hydrolysis of Cellulosic Agricultural Wastes With A Cellulase-Producing Streptomyces For Bioethanol ProductionDocument7 pagesPretreatment and Hydrolysis of Cellulosic Agricultural Wastes With A Cellulase-Producing Streptomyces For Bioethanol ProductionAndré Felipe SilvaNo ratings yet
- Silva 2012Document7 pagesSilva 2012Brian Oro BeltránNo ratings yet
- PHB Process 1998Document10 pagesPHB Process 1998mtlopezNo ratings yet
- 1 s2.0 S1517838217301193 MainDocument7 pages1 s2.0 S1517838217301193 Main7fkqx7zh7pNo ratings yet
- Ethanol Production From WheyDocument4 pagesEthanol Production From WheySalman Zafar100% (1)
- Separation and Purification TechnologyDocument7 pagesSeparation and Purification TechnologyIka SulistyaningtiyasNo ratings yet
- Rhodosporidium Toruloides Y4 in Fed-Batch Culture: High-Density Cultivation of Oleaginous YeastDocument6 pagesRhodosporidium Toruloides Y4 in Fed-Batch Culture: High-Density Cultivation of Oleaginous YeastAnderlone SidabutarNo ratings yet
- Antonopoulou 2016Document41 pagesAntonopoulou 2016Julio MoralesNo ratings yet
- The Exometabolome of Clostridium Thermocellum Reveals Overflow Metabolism at High Cellulose LoadingDocument11 pagesThe Exometabolome of Clostridium Thermocellum Reveals Overflow Metabolism at High Cellulose LoadingMaruf MuhammadNo ratings yet
- A Comparison of The Production of Ethanol Between Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation Using UnpretDocument9 pagesA Comparison of The Production of Ethanol Between Simultaneous Saccharification and Fermentation and Separate Hydrolysis and Fermentation Using Unpretat6315763No ratings yet
- Batch Electrodialysis of Lactic Acid Obtained From LAB FermentationDocument6 pagesBatch Electrodialysis of Lactic Acid Obtained From LAB FermentationkhairulNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese WheyDocument13 pagesFermentation of Lactose To Ethanol in Cheese WheyVitor ArsenioNo ratings yet
- F1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsDocument9 pagesF1C121039 - Finkyasti Nur Adman - Tugas Bahasa Inggris SainsFinkyasti Nur AdmanNo ratings yet
- General Process For The Preparation of Single Cell ProteinsDocument19 pagesGeneral Process For The Preparation of Single Cell ProteinsNikita BatraNo ratings yet
- Fermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColiDocument12 pagesFermentation of Lactose To Ethanol in Cheese Whey Permeate and Concentrated Permeate by Engineered Escherichia ColijoelfloresanchezNo ratings yet
- 1 s2.0 S0306261912004266 Main.41Document6 pages1 s2.0 S0306261912004266 Main.41luuphuongNo ratings yet
- Lactosuero 9 NoDocument24 pagesLactosuero 9 NoMarco Antonio Salcedo HinojosaNo ratings yet
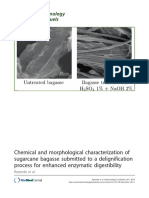
- Chemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityDocument19 pagesChemical and Morphological Characterization of Sugarcane Bagasse Submitted To A Delignification Process For Enhanced Enzymatic DigestibilityTabiita Huaman NorabuenaNo ratings yet
- Biomass and Bioenergy: Research PaperDocument10 pagesBiomass and Bioenergy: Research PaperPaulina Lpz JrzNo ratings yet
- The Potential of Microbial ProcessesDocument18 pagesThe Potential of Microbial ProcessesMichelli SantarelliNo ratings yet
- Economical Feasibility Evaluation of An Ethanol Injection Liposome Production PlantDocument6 pagesEconomical Feasibility Evaluation of An Ethanol Injection Liposome Production Plantraghu_iictNo ratings yet
- Methods For Pretreatment of Lignocellulosic Biomass For Efficient Hydrolysis and Biofuel ProductionDocument18 pagesMethods For Pretreatment of Lignocellulosic Biomass For Efficient Hydrolysis and Biofuel ProductionFrank O'cengNo ratings yet
- Effect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisDocument13 pagesEffect of Dilute Acid Pretreatment of Different Energy Crops On Their Enzymatic HydrolysisProAkademiaNo ratings yet
- Auto-Induction Screening Protocol For Ranking Clonal Libraries of Pichia Pastoris Mut StrainsDocument14 pagesAuto-Induction Screening Protocol For Ranking Clonal Libraries of Pichia Pastoris Mut StrainsAvijeet SinghNo ratings yet
- Fuel Processing Technology: Chun Sheng Goh, Hui Teng Tan, Keat Teong Lee, Abdul Rahman MohamedDocument6 pagesFuel Processing Technology: Chun Sheng Goh, Hui Teng Tan, Keat Teong Lee, Abdul Rahman MohamedHadi AmirkhaniNo ratings yet
- J Biombioe 2010 02 003Document7 pagesJ Biombioe 2010 02 003ABDUL ZIRAY ARIFINNo ratings yet
- Repeated Fed-Batch Lactic Acid Production in A Packed Bed-Stirred Fermentor System Using A PH Feedback Feeding MethodDocument7 pagesRepeated Fed-Batch Lactic Acid Production in A Packed Bed-Stirred Fermentor System Using A PH Feedback Feeding MethodBeeviFathimaNo ratings yet
- Levaduras OleaginosasDocument3 pagesLevaduras OleaginosasChucho LópezNo ratings yet
- Hormesis Effects of Phosphorus On The Viability of Chlorella Regularis Cells Under Nitrogen LimitationDocument9 pagesHormesis Effects of Phosphorus On The Viability of Chlorella Regularis Cells Under Nitrogen LimitationClaudia UngureanuNo ratings yet
- Potential and Assessment of Lactic Acid Production and Isolation - A ReviewDocument34 pagesPotential and Assessment of Lactic Acid Production and Isolation - A ReviewRenata CamilaNo ratings yet
- Jurnal 13 - SkripsiDocument17 pagesJurnal 13 - SkripsiBagusSatriyaNo ratings yet
- Oonkhanond 2017Document37 pagesOonkhanond 2017Gebby FebriliaNo ratings yet
- Evaluation of Different Sludges From WWT20151027-2587-1pylawj PDFDocument10 pagesEvaluation of Different Sludges From WWT20151027-2587-1pylawj PDFBrandeice BarrettNo ratings yet
- Acid and Alkaline Hydrolysis Technologies For Bioethanol Production: An OverviewDocument14 pagesAcid and Alkaline Hydrolysis Technologies For Bioethanol Production: An Overvieweychezdy07No ratings yet
- 1 s2.0 S0360319922042458 MainDocument11 pages1 s2.0 S0360319922042458 Mainranjani093No ratings yet
- Biogas Potential From Anaerobic Co-Digestion of Faecal Sludge With Food Waste and Garden WasteDocument9 pagesBiogas Potential From Anaerobic Co-Digestion of Faecal Sludge With Food Waste and Garden WasteGreen MyanmarNo ratings yet
- Improvements in Titer, Productivity, and Yield Using Solka-Floc For Cellulase ProductionDocument16 pagesImprovements in Titer, Productivity, and Yield Using Solka-Floc For Cellulase Productionguime27No ratings yet
- Evaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationDocument10 pagesEvaluation of Support Matrices For Immobilization of Anaerobic Consortia For e Cient Carbon Cycling in Waste RegenerationGregorius BudiantoNo ratings yet
- KrahmerDocument11 pagesKrahmermndrawNo ratings yet
- Improving Biogas Production From Wheat Plant Using Alkaline PretreatmentDocument6 pagesImproving Biogas Production From Wheat Plant Using Alkaline PretreatmentMaryam TaherdanakNo ratings yet
- Fungi and Lignocellulosic BiomassFrom EverandFungi and Lignocellulosic BiomassChristian P KubicekNo ratings yet
- PDC Lecture Notes - Block Diagram Simplification 2018Document11 pagesPDC Lecture Notes - Block Diagram Simplification 2018R-A Pascual100% (1)
- PDC Lecture Notes 7 - Stability of Closed-Loop Systems 2018Document56 pagesPDC Lecture Notes 7 - Stability of Closed-Loop Systems 2018R-A PascualNo ratings yet
- Fund Chem Reax Eng CH 1Document52 pagesFund Chem Reax Eng CH 1R-A PascualNo ratings yet
- A History of The Pharmaceutical IndustryDocument17 pagesA History of The Pharmaceutical IndustryR-A PascualNo ratings yet
- STP965 Eb.1415051 1 PDFDocument447 pagesSTP965 Eb.1415051 1 PDFpaolaNo ratings yet
- 03 120000 4800000441 Boc Civ Mat 000026 - 01Document56 pages03 120000 4800000441 Boc Civ Mat 000026 - 01Yasir Nisar SyedNo ratings yet
- A Study of Ziegler-Natta Propylene Polymerization Catalysts by Spectroscopic MethodsDocument32 pagesA Study of Ziegler-Natta Propylene Polymerization Catalysts by Spectroscopic Methodsvinit kumarNo ratings yet
- Fanzart Catalogue 2022 v1Document92 pagesFanzart Catalogue 2022 v1Naitik RaoNo ratings yet
- Astm F 106 - 00 PDFDocument6 pagesAstm F 106 - 00 PDFo_l_0No ratings yet
- Checkpoint Chembassador Chapter 1Document4 pagesCheckpoint Chembassador Chapter 1ALIS SUHAIRIN BT ABD GHANI BMNo ratings yet
- Termometro Digital de Oido GracoDocument40 pagesTermometro Digital de Oido GracoPaúl AliNo ratings yet
- Valves and Seats Weir SPM 2 0Document2 pagesValves and Seats Weir SPM 2 0pabloasotoNo ratings yet
- Foam Types TermpaperDocument6 pagesFoam Types TermpaperRolaxNo ratings yet
- Peroxidase Activity of Hemoglobin - Haptoglobin ComplexesDocument14 pagesPeroxidase Activity of Hemoglobin - Haptoglobin ComplexesJoanne MachadoNo ratings yet
- Syllabus JKSSB Forest Guard Forest DepartmentDocument4 pagesSyllabus JKSSB Forest Guard Forest Departmentanil choudharyNo ratings yet
- Elements and Compound Grade 7Document4 pagesElements and Compound Grade 7jingvillaruelNo ratings yet
- Agilent 6890N Network Gas Chromatograph: Data SheetDocument6 pagesAgilent 6890N Network Gas Chromatograph: Data SheetAshraf ahmedNo ratings yet
- BP - Electrical System With Extreme ConditionsDocument17 pagesBP - Electrical System With Extreme Conditions----No ratings yet
- Chemat, F Dan Giancarlo, C. 2013.Microwave-Assisted Extraction For Bioactive Compounds - London. SpringerDocument248 pagesChemat, F Dan Giancarlo, C. 2013.Microwave-Assisted Extraction For Bioactive Compounds - London. SpringerAsri Sofiana MarwahNo ratings yet
- A Contemporary Overview of Silicon Availability in Agricultural SoilsDocument14 pagesA Contemporary Overview of Silicon Availability in Agricultural SoilsIqbal HussainNo ratings yet
- NFPA 20 5 FinalDocument13 pagesNFPA 20 5 FinalMohammed Yasser100% (1)
- Zaki2018 - Bottled WatreDocument9 pagesZaki2018 - Bottled WatreJosé CâmaraNo ratings yet
- StructuralDocument40 pagesStructuralStephanie Joy Delos Reyes67% (3)
- Manual Hydraulic Knockout SetDocument39 pagesManual Hydraulic Knockout SetJuan Manuel LaraNo ratings yet
- Daftar Obat BaksosDocument4 pagesDaftar Obat Baksosandi linaNo ratings yet
- Fyp1 Group Jan 2017 Rev1 - Ipm & PicDocument15 pagesFyp1 Group Jan 2017 Rev1 - Ipm & PicMuhdHishamuddinNo ratings yet
- Manual Sample Release Reagent S1014E RUO 20200623Document1 pageManual Sample Release Reagent S1014E RUO 20200623yonasNo ratings yet