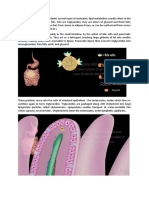
You might also like
- Lipid Profile in AnemiaDocument132 pagesLipid Profile in AnemiaBharath PadmashaliNo ratings yet
- Ninja - Antihyperlipidemics PDFDocument3 pagesNinja - Antihyperlipidemics PDFErica Hyeyeon Lee100% (1)
- LipidsDocument60 pagesLipidsEuneun Bustamante100% (1)
- Lipids Digestion and AbsorptionDocument34 pagesLipids Digestion and Absorptionglenn johnstonNo ratings yet
- CholestrolDocument6 pagesCholestrolprince_woondNo ratings yet
- Chemistry of Fats & OilsDocument38 pagesChemistry of Fats & OilsSubhajit SarkarNo ratings yet
- Lipid MetabolismDocument21 pagesLipid MetabolismTooba GhummanNo ratings yet
- Chemistry of LipidsDocument123 pagesChemistry of LipidsLyra Get100% (1)
- General Biology 1 Q1 Week 7 BiomoleculesDocument5 pagesGeneral Biology 1 Q1 Week 7 BiomoleculesJohn Brylle UrsuaNo ratings yet
- General Biology 1 Q1 Week 7 BiomoleculesDocument5 pagesGeneral Biology 1 Q1 Week 7 BiomoleculesJohn Brylle UrsuaNo ratings yet
- Anexo 2 - Ejercicio 3 - Artículo 2Document12 pagesAnexo 2 - Ejercicio 3 - Artículo 2RocioNo ratings yet
- Hur 2007 PDFDocument9 pagesHur 2007 PDFFábio SilvaNo ratings yet
- Biokimia LipidDocument6 pagesBiokimia Lipiddimensi warnaNo ratings yet
- Atherosclerosis: Reporter: Collera, Charissa Constantino, Venice Clemena, Adnan Coronel, Romeo Cordova, KarlaDocument73 pagesAtherosclerosis: Reporter: Collera, Charissa Constantino, Venice Clemena, Adnan Coronel, Romeo Cordova, Karlaprecious_bustosNo ratings yet
- Bile Acids Bile Acids (Bile Salts) Are Polar Derivatives of Cholesterol. They Are Formed in TheDocument9 pagesBile Acids Bile Acids (Bile Salts) Are Polar Derivatives of Cholesterol. They Are Formed in TheayshmoNo ratings yet
- Metabolism of LipidsDocument98 pagesMetabolism of LipidsLyra GetNo ratings yet
- Lipid Metabolism: Sri Suciati Ningsih FK Uhamka 11 Maret 2020Document39 pagesLipid Metabolism: Sri Suciati Ningsih FK Uhamka 11 Maret 2020annisaaislamNo ratings yet
- Lipid Metabolism - (V) - Cholesterol and LipoproteinsDocument35 pagesLipid Metabolism - (V) - Cholesterol and Lipoproteinslightning proNo ratings yet
- Lipids NotesDocument4 pagesLipids NoteskatNo ratings yet
- Lipoprotein - WikipediaDocument38 pagesLipoprotein - WikipediaTejaswiNo ratings yet
- Approval SheetDocument24 pagesApproval SheetIntan AnggrainiNo ratings yet
- Digestion of Lipids. Transport Forms of LipidsDocument91 pagesDigestion of Lipids. Transport Forms of LipidsAhmed FtsNo ratings yet
- Metabolisme Kolesterol, Lipoprotein Dan ApolipoproteinDocument32 pagesMetabolisme Kolesterol, Lipoprotein Dan ApolipoproteinAlmira Ulfa Utari NasutionNo ratings yet
- BCH 206 CholesterolDocument3 pagesBCH 206 CholesterolNewtonNo ratings yet
- Cholesterol Metabolism: Qassim University College of Medicine Phase II Year II Cardiovascular System BlockDocument23 pagesCholesterol Metabolism: Qassim University College of Medicine Phase II Year II Cardiovascular System BlockichaNo ratings yet
- VojoyDocument7 pagesVojoyFerds SalvatierraNo ratings yet
- CholesterolDocument12 pagesCholesterolCrina LupuNo ratings yet
- Steroid HormonesDocument16 pagesSteroid HormonesRishiNo ratings yet
- L06 LipoproteinsDocument23 pagesL06 LipoproteinsAbdullahi NorNo ratings yet
- Cholesterol: Synthesis and MetabolismDocument45 pagesCholesterol: Synthesis and MetabolismSoffa ShmuelNo ratings yet
- Pre Test: Prepared By: Patricia PradoDocument51 pagesPre Test: Prepared By: Patricia PradoLovely AlosadoNo ratings yet
- Tsi New-1Document8 pagesTsi New-1Markos sitotawNo ratings yet
- Bio 6Document10 pagesBio 6Rose Marie AgustinNo ratings yet
- Biological Molecules: Pentoses, Those With Six Carbons As Hexoses, and Those With Seven Carbons As Heptoses. ExamplesDocument31 pagesBiological Molecules: Pentoses, Those With Six Carbons As Hexoses, and Those With Seven Carbons As Heptoses. ExamplesSunday FavourNo ratings yet
- Chem 113 Lipid ActivityDocument3 pagesChem 113 Lipid ActivityAlliah MendozaNo ratings yet
- STUDY GUIDE - Lipid and Carbohydrate CombinedDocument5 pagesSTUDY GUIDE - Lipid and Carbohydrate CombinedMaya Abou MradNo ratings yet
- Lec 6 Fed State PDFDocument52 pagesLec 6 Fed State PDFrajeshNo ratings yet
- BiochemieDocument10 pagesBiochemiearian.boostani2002No ratings yet
- Cell Bio Library WorkDocument5 pagesCell Bio Library WorkJP SilvaNo ratings yet
- Assignment of Biochemistry 2Document9 pagesAssignment of Biochemistry 2sooraj kumarNo ratings yet
- OliveOil Fact Sheet 08Document5 pagesOliveOil Fact Sheet 08vksk1951No ratings yet
- OliveOil Fact Sheet 08Document5 pagesOliveOil Fact Sheet 08vksk1951No ratings yet
- PARAS, Mark Heinrich B. - Exercise 3 - Lecture Biochemistry CHEM 40 BSN 1EDocument4 pagesPARAS, Mark Heinrich B. - Exercise 3 - Lecture Biochemistry CHEM 40 BSN 1EMark Heinrich B. ParasNo ratings yet
- 5.1 Lipids MetabolismDocument31 pages5.1 Lipids MetabolismAbdallah AlasalNo ratings yet
- Module 11 Lipids 1Document10 pagesModule 11 Lipids 1cassseeeyyyNo ratings yet
- Tsi NewDocument7 pagesTsi NewMarkos sitotawNo ratings yet
- Biochemistry - Fundamentals of Energy Exchange MSCDocument33 pagesBiochemistry - Fundamentals of Energy Exchange MSCmariam1.ashrafNo ratings yet
- Cardiology Update:: LDL Reduction and HDL Management 2012Document8 pagesCardiology Update:: LDL Reduction and HDL Management 2012senaringNo ratings yet
- CarbohydratesDocument6 pagesCarbohydratesGEM MIRASOLNo ratings yet
- BiomoleculesDocument11 pagesBiomoleculesAdul basit mughalNo ratings yet
- Purification of LipidDocument13 pagesPurification of LipidNurakmal SyuhAdaNo ratings yet
- MODULE - FINALS - Biochem - 1Document5 pagesMODULE - FINALS - Biochem - 1Cyril CauilanNo ratings yet
- Balancing Cholesterol Synthesis and AbsorptionDocument5 pagesBalancing Cholesterol Synthesis and AbsorptionPoppy DharsanaNo ratings yet
- 1.2-1.11 Carbohydrates and ProteinsDocument7 pages1.2-1.11 Carbohydrates and ProteinsbritNo ratings yet
- Lipid Metabolism - WikipediaDocument31 pagesLipid Metabolism - WikipediaTomiwa AdeshinaNo ratings yet
- Metabolism of LipidsDocument96 pagesMetabolism of LipidsLyra GetNo ratings yet
- 9-23-2010 BiomoleculesDocument7 pages9-23-2010 BiomoleculesalbornthealchemistNo ratings yet
- 1.overview of MetabolismDocument6 pages1.overview of Metabolismقتيبه خالد دحام خلفNo ratings yet
- Narasi Presentasi Bagian Mekanisme Lipid (B11)Document4 pagesNarasi Presentasi Bagian Mekanisme Lipid (B11)Lana WintersNo ratings yet
- Carbon: BiomoleculesDocument16 pagesCarbon: BiomoleculesJoseph GuerreroNo ratings yet
- Tenp DPT Biochemistry: DR Pius KiptemburDocument13 pagesTenp DPT Biochemistry: DR Pius KiptemburGerald Limo Arap ChebiiNo ratings yet
- Lipid MetabolismDocument52 pagesLipid MetabolismDr.P.NatarajanNo ratings yet
- Disorders of Lipid Metabolism Full Article Tables FiguresDocument32 pagesDisorders of Lipid Metabolism Full Article Tables Figureshasnawati100% (1)
- Cholesterol Synthesis PathwayDocument16 pagesCholesterol Synthesis PathwayaddyNo ratings yet
- The Truth About Dietary Cholesterol - Based On The Teachings Of Dr. Andrew Huberman: Separating Truth From MythsFrom EverandThe Truth About Dietary Cholesterol - Based On The Teachings Of Dr. Andrew Huberman: Separating Truth From MythsNo ratings yet
- Class 1 Reading, Food From The EarthDocument3 pagesClass 1 Reading, Food From The EarthDiego Armando Quevedo LuquinNo ratings yet
- App G4U (A1 - Test Unit 1, 2 & 3)Document9 pagesApp G4U (A1 - Test Unit 1, 2 & 3)Diego Armando Quevedo LuquinNo ratings yet
- Butirometro para Leche FT PDFDocument15 pagesButirometro para Leche FT PDFAlejandro Lara ResendízNo ratings yet
- PT 01Document13 pagesPT 01Dennis LimNo ratings yet
- Part 05 ADocument25 pagesPart 05 ADiego Armando Quevedo LuquinNo ratings yet
- FOLIODocument1 pageFOLIODiego Armando Quevedo LuquinNo ratings yet
- WinterizationDocument11 pagesWinterizationmsarriatorresNo ratings yet
- Classification of LipidsDocument18 pagesClassification of LipidsD Kaur100% (1)
- PN-1150653405 OCHOL Sellsheet DISTDocument4 pagesPN-1150653405 OCHOL Sellsheet DISTMarin MarianNo ratings yet
- Is There Still A Future For Hydrogenated Vegetable Oils? : An Philippaerts, Pierre A. Jacobs, and Bert F. SelsDocument8 pagesIs There Still A Future For Hydrogenated Vegetable Oils? : An Philippaerts, Pierre A. Jacobs, and Bert F. SelsamiraNo ratings yet
- Regulation of Cholesterol SynthesisDocument20 pagesRegulation of Cholesterol SynthesisHusnaNo ratings yet
- R. K. Life Services Private Limited (Department of Laboratory Medicine)Document1 pageR. K. Life Services Private Limited (Department of Laboratory Medicine)Aniruddha ChatterjeeNo ratings yet
- Carnauba WaxDocument3 pagesCarnauba WaxTenesaca OscarNo ratings yet
- Media Medika IndonesianaDocument8 pagesMedia Medika IndonesianaMuhammad Hamzah AsadullahNo ratings yet
- 1981 Composition ScholfieldDocument4 pages1981 Composition ScholfieldAngela María García MoraNo ratings yet
- Paket TerbaruDocument8 pagesPaket TerbaruAmmey JoizeNo ratings yet
- Lipid Chemistry 1Document102 pagesLipid Chemistry 1queenNo ratings yet
- Essential Fatty Acids in Clinical Dermatology PDFDocument9 pagesEssential Fatty Acids in Clinical Dermatology PDFSonia CaruntuNo ratings yet
- JURNALDocument9 pagesJURNALAnnisa HasanahNo ratings yet
- Analytic HDL OdfDocument15 pagesAnalytic HDL OdfanggaririnNo ratings yet
- Pengaruh Pemberian Minyak Kanola Terhadap Gambaran Histopatologik Aorta Dan Kadar Kolesterol Tikus Wistar Dengan Diet Tinggi LemakDocument4 pagesPengaruh Pemberian Minyak Kanola Terhadap Gambaran Histopatologik Aorta Dan Kadar Kolesterol Tikus Wistar Dengan Diet Tinggi LemakDedi Satria WahyuNo ratings yet
- Lipid PDFDocument5 pagesLipid PDFKhổng Tiến ĐạtNo ratings yet
- Accepted Manuscript: Epilepsy ResearchDocument14 pagesAccepted Manuscript: Epilepsy ResearchSyedNo ratings yet
- DyslipidemiaDocument50 pagesDyslipidemiaTitin Tria UtamiNo ratings yet
- Relacion Colesterol Total A HDL y Colesterol No HD PDFDocument9 pagesRelacion Colesterol Total A HDL y Colesterol No HD PDFJoseAbdalaNo ratings yet
- LipidsDocument53 pagesLipidsGlen Mangali100% (1)
- 0043 PDFDocument12 pages0043 PDFZuhdan AnnajmutstsaqibNo ratings yet
- Other Carbohydrates - Paper - Ethanol Fats and Oils: Q. Laurence Salmon BarrantesDocument32 pagesOther Carbohydrates - Paper - Ethanol Fats and Oils: Q. Laurence Salmon BarrantesCRISTHIANNo ratings yet
- Chapter 08 LipidDocument10 pagesChapter 08 Lipid楊畯凱No ratings yet
- Digestion and Absorption of FatsDocument4 pagesDigestion and Absorption of Fatsceer1991No ratings yet
- P.B. TELEFO, Ph.D. Senior Lecturer of Biochemistry University of Dschang, CameroonDocument59 pagesP.B. TELEFO, Ph.D. Senior Lecturer of Biochemistry University of Dschang, CameroonTabe AndersonNo ratings yet