You might also like
- Python Cheat SheetDocument14 pagesPython Cheat Sheetgr8ganesh4495100% (2)
- Microbiology: a QuickStudy Laminated 6-Page Reference GuideFrom EverandMicrobiology: a QuickStudy Laminated 6-Page Reference GuideNo ratings yet
- BMI1014: Chapter 2 - Bacteria-Morphology and ClassificationDocument6 pagesBMI1014: Chapter 2 - Bacteria-Morphology and ClassificationAkmal Adib FadzilNo ratings yet
- Guyton Hall Medical Physiology 13th Test BankDocument5 pagesGuyton Hall Medical Physiology 13th Test BankOsman Nazir33% (3)
- Advanced Hindi Grammar Table of ContentsDocument156 pagesAdvanced Hindi Grammar Table of ContentsSwetha RameshNo ratings yet
- World Top 2% Scientists From India From Various FieldsDocument25 pagesWorld Top 2% Scientists From India From Various FieldsThe Indian Express100% (14)
- World Top 2% Scientists From India From Various FieldsDocument25 pagesWorld Top 2% Scientists From India From Various FieldsThe Indian Express100% (14)
- World Top 2% Scientists From India From Various FieldsDocument25 pagesWorld Top 2% Scientists From India From Various FieldsThe Indian Express100% (14)
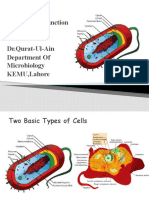
- Bacterial Cell Structure FunctionDocument35 pagesBacterial Cell Structure FunctionItzz MilzzNo ratings yet
- Concepts of BiologyDocument615 pagesConcepts of BiologyGrant M EsteppNo ratings yet
- Morphology of BacteriaDocument44 pagesMorphology of BacteriaDr Harender SimarNo ratings yet
- Prokaryotic CellDocument29 pagesProkaryotic CellSeshime Thyrone DavidsonNo ratings yet
- NUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityDocument4 pagesNUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityPrancheska Abigayle Peneyra SantiagoNo ratings yet
- Lecture 3 - Bacterial Anatomy and PhysiologyDocument26 pagesLecture 3 - Bacterial Anatomy and Physiologyapi-370335280% (5)
- Cells Questions and VocabDocument12 pagesCells Questions and VocabFanna Sharma100% (1)
- DETAILED LP Distance Vs DisplacementDocument8 pagesDETAILED LP Distance Vs DisplacementDiane Marr Nicolas DencioNo ratings yet
- Bacterial Structure, Growth, and Metabolism ExplainedDocument77 pagesBacterial Structure, Growth, and Metabolism ExplainedNelle ReneiNo ratings yet
- Full Download Brock Biology of Microorganisms 15th Edition Madigan Test BankDocument35 pagesFull Download Brock Biology of Microorganisms 15th Edition Madigan Test Bankbeizatikeorar100% (20)
- Meningitidis), or More Than Once To Produce A Chain (Streptococcus Pyogenes), Divides Regularly in Two Planes at RightDocument14 pagesMeningitidis), or More Than Once To Produce A Chain (Streptococcus Pyogenes), Divides Regularly in Two Planes at RightMichael Vincent Pizarro BarbaNo ratings yet
- BITS Pilani Morphology and Structure of BacteriaDocument25 pagesBITS Pilani Morphology and Structure of Bacteriasid011No ratings yet
- Micro para 1Document4 pagesMicro para 1Reselle EspirituNo ratings yet
- Bacterial Morphology and Cell StructureDocument14 pagesBacterial Morphology and Cell StructureJe KirsteneNo ratings yet
- Part 10 ProkaryoticDocument54 pagesPart 10 ProkaryoticLê Thanh HằngNo ratings yet
- Bacterial Cell Structures - Dr NBora_19112018 (1)Document35 pagesBacterial Cell Structures - Dr NBora_19112018 (1)jeozbapqfpugltpsrbNo ratings yet
- Glycocalyx and Cell Wall of BacteriaDocument3 pagesGlycocalyx and Cell Wall of BacteriaHrushikesh KulkarniNo ratings yet
- Microbial Taxonomy - System That Involves in The Organization, Classification, Naming orDocument5 pagesMicrobial Taxonomy - System That Involves in The Organization, Classification, Naming orRedelle Mae NiniNo ratings yet
- bacterial notes 2022Document5 pagesbacterial notes 2022Banji MaikaNo ratings yet
- Mod 2Document24 pagesMod 2The GreatNo ratings yet
- TUTORIAL 3 MicrobiologyDocument8 pagesTUTORIAL 3 MicrobiologyJOSEPH TING FU KIONGNo ratings yet
- Prokaryotic Cells: Cell Wall: Made of A Murein (Not Cellulose), Which Is A Glycoprotein or PeptidoglycanDocument3 pagesProkaryotic Cells: Cell Wall: Made of A Murein (Not Cellulose), Which Is A Glycoprotein or Peptidoglycanserah1024No ratings yet
- 1 - Introduction To Medical MicrobiologyDocument63 pages1 - Introduction To Medical MicrobiologyAhmed MohamedNo ratings yet
- Procaryotic Cell ArchitectureDocument18 pagesProcaryotic Cell ArchitectureWindi Dawn SallevaNo ratings yet
- Prokaryotic Cell Structure Function LectureDocument39 pagesProkaryotic Cell Structure Function LectureMisbah ShabbirNo ratings yet
- BDS Year 2 Lecture on Prokaryotic and Eukaryotic Cell StructuresDocument46 pagesBDS Year 2 Lecture on Prokaryotic and Eukaryotic Cell StructuresNarmathaa ThevarNo ratings yet
- Structure of Bact P.E.2018Document60 pagesStructure of Bact P.E.2018Ludmila BuzurniiNo ratings yet
- DM-1 - Lesson 11Document10 pagesDM-1 - Lesson 11Amit GodaraNo ratings yet
- Bacteria: Cell Walls: Structure of PeptidoglycanDocument14 pagesBacteria: Cell Walls: Structure of PeptidoglycanReese SoleilNo ratings yet
- Lecture 1 Bacterial StructureDocument38 pagesLecture 1 Bacterial StructureAyat MostafaNo ratings yet
- Structure/Function of Bacteria: BIOL 2900 - Week 3Document5 pagesStructure/Function of Bacteria: BIOL 2900 - Week 3PaigufanNo ratings yet
- Chapter 8 Points To RememberDocument5 pagesChapter 8 Points To RememberNANDA KISHORE S X A AD : 1 0 6 4 4No ratings yet
- Bacterial Classification and Morphology GuideDocument4 pagesBacterial Classification and Morphology GuideJerson Aizpuro SuplementoNo ratings yet
- Lecture 1 Introduction PDFDocument51 pagesLecture 1 Introduction PDFMomo KimNo ratings yet
- CellDocument16 pagesCellanantchouhdary1709No ratings yet
- Morfology AND Citology of Bacteria: Sidarningsih, Drg.,mkesDocument47 pagesMorfology AND Citology of Bacteria: Sidarningsih, Drg.,mkesDaffa YudhistiraNo ratings yet
- Microbiology Summaries 1-30 ReviewedDocument53 pagesMicrobiology Summaries 1-30 ReviewedyepNo ratings yet
- Bacterial Morphology and StructureDocument68 pagesBacterial Morphology and Structureapi-19916399No ratings yet
- Cell WallDocument1 pageCell WallFrancine ValdezNo ratings yet
- Lecture 2 - Bacterial Cell Structure and FunctionDocument90 pagesLecture 2 - Bacterial Cell Structure and FunctionOdurNo ratings yet
- Nama: Muhammad Hafiz NIM: 1710814110016 Prodi: S1 Teknik Kimia Matkul: Mikrobiologi IndustriDocument7 pagesNama: Muhammad Hafiz NIM: 1710814110016 Prodi: S1 Teknik Kimia Matkul: Mikrobiologi IndustriHafiz MuhammadNo ratings yet
- Bacterial Cell Structure and ReproductionDocument10 pagesBacterial Cell Structure and Reproductionمنتظر علي حسين A2No ratings yet
- Bacterial Anatomy BScNursingDocument31 pagesBacterial Anatomy BScNursingtshering chodenNo ratings yet
- Structure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovDocument54 pagesStructure & Functions of Prokaryotic Cell. Bacteriophages.: Lecturer: - Sergei V. RedkozubovJustineNo ratings yet
- 11 Biology Revision Study Material Chapter 8Document8 pages11 Biology Revision Study Material Chapter 8Saurav SoniNo ratings yet
- 5 Prokaryatic Structure and Function 5Document31 pages5 Prokaryatic Structure and Function 5Sadam IrshadNo ratings yet
- Julie Bui - Bacterial Structure FunctionDocument2 pagesJulie Bui - Bacterial Structure Functionapi-522650514No ratings yet
- StructureDocument15 pagesStructurefatimasanusimorikiNo ratings yet
- Prokaryotic Cell Structure and FunctionsDocument39 pagesProkaryotic Cell Structure and Functionsabida waryamNo ratings yet
- Bacterial Cell Wall TypesDocument7 pagesBacterial Cell Wall Typess.khan9211rediffmail.comNo ratings yet
- The Structure of Bacterial CellDocument49 pagesThe Structure of Bacterial CellanaNo ratings yet
- The Physiology of Prokaryotic Cell: Understanding the Smallest Form of LifeDocument22 pagesThe Physiology of Prokaryotic Cell: Understanding the Smallest Form of LifeAnnysaNurmalaSariNo ratings yet
- Medical Microbiology Course Bacterial Cell StructureDocument5 pagesMedical Microbiology Course Bacterial Cell StructureariffdrNo ratings yet
- For Ntse CellDocument73 pagesFor Ntse CellPrakhar RajNo ratings yet
- Micro Lecture 2: Structure and Characteristics of Bacterial CellsDocument78 pagesMicro Lecture 2: Structure and Characteristics of Bacterial CellsJaellah MatawaNo ratings yet
- Functional Anatomy of Prokaryotes and EukaryotesDocument6 pagesFunctional Anatomy of Prokaryotes and EukaryotesMollyNo ratings yet
- Bacterial Cell StructureDocument9 pagesBacterial Cell StructureManWol JangNo ratings yet
- Bacterial Cell Structure and MorphologyDocument45 pagesBacterial Cell Structure and Morphologyauguz21acenaNo ratings yet
- Chapter 3 Micropara (Outline)Document5 pagesChapter 3 Micropara (Outline)Jezrylle BalaongNo ratings yet
- Chapter 1,2,3 Micro PaperDocument9 pagesChapter 1,2,3 Micro Papermenamamdouh20No ratings yet
- Infectious Disease NotesDocument43 pagesInfectious Disease NotessaguNo ratings yet
- Bacterial MorphologyDocument75 pagesBacterial MorphologyJoan MelendresNo ratings yet
- Enzyme Notes - SathyanarayanaDocument32 pagesEnzyme Notes - SathyanarayanaSwetha RameshNo ratings yet
- PG Curriculum 2019 PDFDocument413 pagesPG Curriculum 2019 PDFsangeerth k babuNo ratings yet
- E.Coli Expression SystemsDocument5 pagesE.Coli Expression SystemsSwetha RameshNo ratings yet
- His Tag Purification PDFDocument5 pagesHis Tag Purification PDFSwetha RameshNo ratings yet
- GATE Exams BT Biotechnology Syllabus 2022Document3 pagesGATE Exams BT Biotechnology Syllabus 2022Swetha RameshNo ratings yet
- Zone/Scrutiny Code Details for GATE 2021 ApplicantDocument1 pageZone/Scrutiny Code Details for GATE 2021 ApplicantSwetha RameshNo ratings yet
- The Cartoon Guide To Statistics-3Document8 pagesThe Cartoon Guide To Statistics-3Swetha Ramesh100% (1)
- Cells of Immune SystemDocument14 pagesCells of Immune SystemSwetha RameshNo ratings yet
- Competant Cells and TransformationDocument3 pagesCompetant Cells and TransformationSwetha RameshNo ratings yet
- Lancet Journal On CovidDocument2 pagesLancet Journal On CovidSwetha RameshNo ratings yet
- T CellsDocument11 pagesT CellsSwetha RameshNo ratings yet
- Developing Neem-Based Bioplastic for Food PackagingDocument8 pagesDeveloping Neem-Based Bioplastic for Food PackagingSwetha RameshNo ratings yet
- Appendix 1Document29 pagesAppendix 1Swetha RameshNo ratings yet
- NK and MacrophageDocument10 pagesNK and MacrophageSwetha RameshNo ratings yet
- TamilBrahmins.com Credits PageDocument25 pagesTamilBrahmins.com Credits PageSwetha RameshNo ratings yet
- Lymphoid OrganDocument23 pagesLymphoid OrganSwetha RameshNo ratings yet
- Sterilisation Methods-1 PDFDocument9 pagesSterilisation Methods-1 PDFSwetha RameshNo ratings yet
- Next Step Portal Final CCDocument34 pagesNext Step Portal Final CCSwetha RameshNo ratings yet
- Antimicrobial Activities of Leaf ExtractsDocument7 pagesAntimicrobial Activities of Leaf ExtractsMorgana Romero PalomarNo ratings yet
- Plant Tissue Culture-1Document19 pagesPlant Tissue Culture-1Swetha RameshNo ratings yet
- Effects of Processing & Storage on Orange Juice AromaDocument13 pagesEffects of Processing & Storage on Orange Juice AromaSwetha RameshNo ratings yet
- International WebinarDocument1 pageInternational WebinarSwetha RameshNo ratings yet
- Enzymes Improve Textile Scouring and Dyeing ProcessesDocument2 pagesEnzymes Improve Textile Scouring and Dyeing ProcessesSwetha RameshNo ratings yet
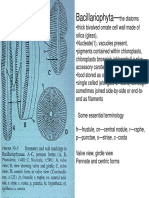
- Bacillariophyta—the Diatoms and Their Unique Silica Cell WallsDocument23 pagesBacillariophyta—the Diatoms and Their Unique Silica Cell WallsSwetha RameshNo ratings yet
- Restriction Mapping - KeyDocument6 pagesRestriction Mapping - KeySwetha RameshNo ratings yet
- 7.1-7.2 Review KeyDocument9 pages7.1-7.2 Review KeyirwindeepsinghNo ratings yet
- Histology Exam I OutlineDocument76 pagesHistology Exam I OutlinePrinceNo ratings yet
- Rampado2022 - Lysis BufferDocument11 pagesRampado2022 - Lysis BufferRamonaTecucianuNo ratings yet
- Active and Passive TransportDocument2 pagesActive and Passive TransportMaricon TubonNo ratings yet
- Animal Cells PDFDocument4 pagesAnimal Cells PDFFalah HabibNo ratings yet
- FOLIO: Biology Form 4 Chapter 2: Cell Structure and OrganisationDocument51 pagesFOLIO: Biology Form 4 Chapter 2: Cell Structure and OrganisationSashimi 刺身 Chebby100% (1)
- 1.1 Basic Principles of Cell InjuryDocument41 pages1.1 Basic Principles of Cell InjuryMUNEZERO Evase0% (1)
- Bio 1 K12Document141 pagesBio 1 K12Donna VallesteroNo ratings yet
- Molecular Physiology of Water BalanceDocument16 pagesMolecular Physiology of Water Balancebiomedikbiokimiausu2014No ratings yet
- A Membranous Sac Which Occupies A Large Part of A Mature Plant CellDocument29 pagesA Membranous Sac Which Occupies A Large Part of A Mature Plant CellintannormalaNo ratings yet
- Farmacologie Intr.Document97 pagesFarmacologie Intr.Sarra GannouniNo ratings yet
- BIOZONE IB Biology 2nd Edition Sample PacketDocument11 pagesBIOZONE IB Biology 2nd Edition Sample Packetvf5ctjknk6No ratings yet
- Volland Et Al 2022Document7 pagesVolland Et Al 2022Juan Sebastian Rodriguez MartinezNo ratings yet
- CELL TRANSPORTPOWEPOINT (Recovered)Document140 pagesCELL TRANSPORTPOWEPOINT (Recovered)ntakirutimana valensNo ratings yet
- Cell Structure and Function RDDocument18 pagesCell Structure and Function RDapi-258400031No ratings yet
- Chapter 3: Compartmentation: Cells and Tissues: Functional Compartments of The BodyDocument10 pagesChapter 3: Compartmentation: Cells and Tissues: Functional Compartments of The BodyShivani RailowalNo ratings yet
- Cell MembraneDocument4 pagesCell MembraneNoraNo ratings yet
- Cell Membrane: Fourth TermDocument10 pagesCell Membrane: Fourth TermThe Troll OMG 2 The Troll OMG 2No ratings yet
- Pulsed Electric Field Processing of FoodsDocument30 pagesPulsed Electric Field Processing of FoodsMohammad Ishfaq BhatNo ratings yet
- Pharmacokinetics in Drug Discovery: A Review of Concepts, Tools, and StrategiesDocument37 pagesPharmacokinetics in Drug Discovery: A Review of Concepts, Tools, and StrategieszahidNo ratings yet
- Materi Kuliah BHL (Moluska - Jurnal - 3) - 2021 DaringDocument20 pagesMateri Kuliah BHL (Moluska - Jurnal - 3) - 2021 DaringAlifia Nabilla PutriNo ratings yet
- BIO101 Handouts by Hanzla Full 130 LessonsDocument225 pagesBIO101 Handouts by Hanzla Full 130 LessonsZeeshan NazarNo ratings yet
- Archaea 2016 PDFDocument73 pagesArchaea 2016 PDFDr. Farokh Rokhbakhsh-Zamin50% (2)
- Biophysics-Lectures-2012-3 (Physiology) PDFDocument107 pagesBiophysics-Lectures-2012-3 (Physiology) PDFNedelcuGeorgeNo ratings yet
- Chapter 7Document13 pagesChapter 7Apryll DarlineNo ratings yet