You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5796)
- Waste Management Plan-1Document16 pagesWaste Management Plan-1usama basionyNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Engine WDshowContent DocumeDocument17 pagesEngine WDshowContent DocumeJoe Anrean JoeNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- 3 - PorphyriasDocument14 pages3 - PorphyriasHamzehNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- MODULE 7-11 Notes PrefiDocument7 pagesMODULE 7-11 Notes PrefiPASCUAL, ALJON R.No ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Freedom WritersDocument2 pagesFreedom WritersAizellah DietoNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (589)
- Covid-19: Testing TimesDocument2 pagesCovid-19: Testing TimeshgfksqdnhcuiNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Code of Business Conduct For Suppliers To The Coca-Cola CompanyDocument1 pageCode of Business Conduct For Suppliers To The Coca-Cola CompanyMonicaNo ratings yet
- AV SensoresDocument1 pageAV SensoresDiego RestagnoNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Carrot Cake Cupcakes - Sugar Spun RunDocument2 pagesCarrot Cake Cupcakes - Sugar Spun RunSean BermejoNo ratings yet
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- 0568Document77 pages0568selvakrishna0% (1)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Persuasive Essay HomeworkDocument8 pagesPersuasive Essay Homeworkafhbgohob100% (2)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Sepam Range High Impedance Differential ProtectionDocument12 pagesSepam Range High Impedance Differential ProtectionosmpotNo ratings yet
- TransformerDocument52 pagesTransformerorangramaiNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Unit-8 - Assignment-1 - WAGOLL 4Document3 pagesUnit-8 - Assignment-1 - WAGOLL 4Alisha AliNo ratings yet
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- UFFL Company Case StudyDocument6 pagesUFFL Company Case StudyDell CaasieNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Lower GI BleedingDocument40 pagesLower GI BleedingMohammad Firdaus100% (2)
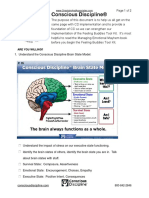
- Conscious Discipline®: Are You Willing?Document2 pagesConscious Discipline®: Are You Willing?Viplav AnandNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- VOIDING TRANSACTIONS in ALLSCRIPTSDocument1 pageVOIDING TRANSACTIONS in ALLSCRIPTSluzmercNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Helical Coil Heat Exchanger Without Agitation (Batch and Continuous) Mas GitoDocument104 pagesHelical Coil Heat Exchanger Without Agitation (Batch and Continuous) Mas GitosehonoNo ratings yet
- South 4Document24 pagesSouth 4LariNo ratings yet
- Pelaez Gabriella M5A1Document3 pagesPelaez Gabriella M5A1Gabriella PelaezNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Blue Mind - The Surprising Science That Shows How Being Near, In, On, or Under Water Can Make You Happier, Healthier, More Connected, and Better at What You Do (PDFDrive)Document246 pagesBlue Mind - The Surprising Science That Shows How Being Near, In, On, or Under Water Can Make You Happier, Healthier, More Connected, and Better at What You Do (PDFDrive)Saddam Akbar100% (5)
- Registration of Business Name-Companies and Allied Matters Act of 1990Document2 pagesRegistration of Business Name-Companies and Allied Matters Act of 1990Osemwengie OsahonNo ratings yet
- Alur WBI Online CrusherDocument7 pagesAlur WBI Online Crusheremping 72No ratings yet
- Stabilization of Soil by Plastic Waste I PDFDocument3 pagesStabilization of Soil by Plastic Waste I PDFDennis Vincent VillarNo ratings yet
- Ground Water Quality Monitoring in A Developing Area Using Water Quality Index and Principal Component Analysis: A Case Study of Visakhapatnam IndiaDocument8 pagesGround Water Quality Monitoring in A Developing Area Using Water Quality Index and Principal Component Analysis: A Case Study of Visakhapatnam IndiaMallikarjuna Rao DuvvadaNo ratings yet
- Experimental Study On The Hardened Properties of Concrete by Using Soft Drink Bottle Caps As Partial Replacement For Coarse AggregatesDocument7 pagesExperimental Study On The Hardened Properties of Concrete by Using Soft Drink Bottle Caps As Partial Replacement For Coarse AggregatesSajjad AhmadNo ratings yet
- ICT - Css 9 Getting To Know Your Computer: Quarter 1Document21 pagesICT - Css 9 Getting To Know Your Computer: Quarter 1Carlota GarciaNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Final Program Book ICBBB 2018Document16 pagesFinal Program Book ICBBB 2018Abida Raza RaoNo ratings yet
- Video Essay Script Draft 2Document2 pagesVideo Essay Script Draft 2api-480313267No ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)