You might also like
- Signal Processing in Auditory Neuroscience: Temporal and Spatial Features of Sound and SpeechFrom EverandSignal Processing in Auditory Neuroscience: Temporal and Spatial Features of Sound and SpeechNo ratings yet
- Elizabeth Lhost and Richard Ashley - Jazz Blues and The Language of Harmony Flexibility in Online Harmonic ProcessingDocument7 pagesElizabeth Lhost and Richard Ashley - Jazz Blues and The Language of Harmony Flexibility in Online Harmonic Processingato.compostoNo ratings yet
- The Role of Syllables in Speech PerceptionDocument7 pagesThe Role of Syllables in Speech PerceptionAhmed S. MubarakNo ratings yet
- Transfer Effects in The Vocal Imitation of Speech and SongDocument18 pagesTransfer Effects in The Vocal Imitation of Speech and SongMonta TupčijenkoNo ratings yet
- Structural 06 Jas Is TDocument11 pagesStructural 06 Jas Is TGreg SurgesNo ratings yet
- 2009 Making Psycholinguistics MusicalDocument8 pages2009 Making Psycholinguistics MusicalAlicia MenesesNo ratings yet
- The Sweet Sounds of Hierarchical Processing in Music and LanguageDocument16 pagesThe Sweet Sounds of Hierarchical Processing in Music and LanguagesertimoneNo ratings yet
- Chiappe & Schmuckler 1997Document6 pagesChiappe & Schmuckler 1997Sepide KNo ratings yet
- Speech Perception: FurtherDocument28 pagesSpeech Perception: FurtherAnastasia NikolaevaNo ratings yet
- The Neural Code ThatDocument2 pagesThe Neural Code ThatchysaNo ratings yet
- A Coarticulatory Path To Sound ChangeDocument38 pagesA Coarticulatory Path To Sound ChangeMarina LacerdaNo ratings yet
- De Leeuw, Andrews Et Al. 2018Document15 pagesDe Leeuw, Andrews Et Al. 2018Nimish AkulaNo ratings yet
- Music, Rhythm, Rise Time Perception and Developmental Dyslexia Perception of Musical Meter Predicts Reading and PhonologyDocument16 pagesMusic, Rhythm, Rise Time Perception and Developmental Dyslexia Perception of Musical Meter Predicts Reading and Phonologybiahonda15No ratings yet
- Rhythmic and Speech Rate Effects in The Perception of Durational CuesDocument21 pagesRhythmic and Speech Rate Effects in The Perception of Durational Cuesdielan wuNo ratings yet
- Dankovicova 2007Document49 pagesDankovicova 2007JEREMY SHANDEL BECERRA DELGADONo ratings yet
- Transfer Effects in The Vocal Imitation of Speech and SongDocument18 pagesTransfer Effects in The Vocal Imitation of Speech and SongMonta TupčijenkoNo ratings yet
- Bidelman Et Al. - 2014 - Coordinated Plasticity in Brainstem and Auditory Cortex MARKUPDocument12 pagesBidelman Et Al. - 2014 - Coordinated Plasticity in Brainstem and Auditory Cortex MARKUPricardo MillaNo ratings yet
- Analyzing Sound Tracings - A Multimodal Approach To Music Information RetrievalDocument6 pagesAnalyzing Sound Tracings - A Multimodal Approach To Music Information RetrievalMiri CastilloNo ratings yet
- Fowler 1994 PDFDocument14 pagesFowler 1994 PDFCami MarquezNo ratings yet
- Imitate or Recall: How Do Musicians Perform Raga Phrases?Document7 pagesImitate or Recall: How Do Musicians Perform Raga Phrases?Ranjan Sengupta100% (1)
- Exploring Cognitive Relations Between Prediction in Language and MusicDocument18 pagesExploring Cognitive Relations Between Prediction in Language and Musicbiahonda15No ratings yet
- Resolving Lexical Ambiguity With Sub-Phonemic Information: I Scream For Ice CreamDocument24 pagesResolving Lexical Ambiguity With Sub-Phonemic Information: I Scream For Ice Creamhala Al-obeideyNo ratings yet
- 10 1111@j 1460-9568 2006 04785 X PDFDocument13 pages10 1111@j 1460-9568 2006 04785 X PDFTlaloc GonzalezNo ratings yet
- Frieler Constructing Jazz Lines 2019Document30 pagesFrieler Constructing Jazz Lines 2019ClaudiaFerri100% (1)
- Memory For Musical Tempo Additional Evidence That Auditory Memory Is AbsoluteDocument9 pagesMemory For Musical Tempo Additional Evidence That Auditory Memory Is AbsoluteLeonardo AguiarNo ratings yet
- The Whole is Not Different From its PartsDocument17 pagesThe Whole is Not Different From its PartsFarlley DerzeNo ratings yet
- Speech Perception NotesDocument13 pagesSpeech Perception NotesMuhammad SanwalNo ratings yet
- Toussaint - The Pairwise Variability Index As A Tool in Musical Rhythm AnalysisDocument8 pagesToussaint - The Pairwise Variability Index As A Tool in Musical Rhythm AnalysisAlessandro Ratoci100% (1)
- Palomary Zatorre Cerebral CortexDocument11 pagesPalomary Zatorre Cerebral Cortexmcarmen rodriguez navasNo ratings yet
- NIH Public Access: Investigation of Musicality in BirdsongDocument27 pagesNIH Public Access: Investigation of Musicality in Birdsonglamaildierica9188No ratings yet
- Psychology of Music: Is Expressive Timing Relational Invariant Under Tempo Transformation?Document11 pagesPsychology of Music: Is Expressive Timing Relational Invariant Under Tempo Transformation?xmlbioxNo ratings yet
- Content 2001 Content2001tnDocument23 pagesContent 2001 Content2001tnAlain ContentNo ratings yet
- Can Nonlinguistic Musical Training Change The Way The Brain Processes Speech? The Expanded OPERA HypothesisDocument11 pagesCan Nonlinguistic Musical Training Change The Way The Brain Processes Speech? The Expanded OPERA HypothesisAna Carolina RibasNo ratings yet
- The Neural Mechanisms of Speech Comprehension: fMRI Studies of Semantic AmbiguityDocument9 pagesThe Neural Mechanisms of Speech Comprehension: fMRI Studies of Semantic AmbiguityyuvharpazNo ratings yet
- Distinct Brain Systems For Processing Concrete and Abstract ConceptsDocument13 pagesDistinct Brain Systems For Processing Concrete and Abstract ConceptstkorotajdracaNo ratings yet
- "Moving To The Beat" Improves Timing Perception: Brief ReportDocument7 pages"Moving To The Beat" Improves Timing Perception: Brief ReportmoiNo ratings yet
- About Sentential ContextDocument21 pagesAbout Sentential Contexthala Al-obeideyNo ratings yet
- An Empirical Comparison of Rhythm in Language and Music: Aniruddh D. Patel, Joseph R. DanieleDocument11 pagesAn Empirical Comparison of Rhythm in Language and Music: Aniruddh D. Patel, Joseph R. DanieleGiacomo Dal PraNo ratings yet
- Bermudez, Zatorre - 2009 - The Absolute Pitch Mind Continues To Reveal ItselfDocument4 pagesBermudez, Zatorre - 2009 - The Absolute Pitch Mind Continues To Reveal ItselfbenqtenNo ratings yet
- Matsunagag, 2015Document13 pagesMatsunagag, 2015Sharon Rozenbaum LagonNo ratings yet
- Dial2019 - Article - LexicalProcessingDependsOnSublDocument18 pagesDial2019 - Article - LexicalProcessingDependsOnSublMarcela MacedoNo ratings yet
- Individual DifferencesDocument10 pagesIndividual DifferencesJuan CastroNo ratings yet
- (Arcara Et Al., 2014) Word Structure and Decomposition Effects in ReadingDocument36 pages(Arcara Et Al., 2014) Word Structure and Decomposition Effects in ReadingLê Huỳnh PhúcNo ratings yet
- Bonatti V Vs C2004 PDFDocument9 pagesBonatti V Vs C2004 PDFJose Luis Verona GomezNo ratings yet
- Cummins 2009 JPhon09Document13 pagesCummins 2009 JPhon09ryuslkaNo ratings yet
- Mannel, 2016Document7 pagesMannel, 2016bb9mw7gr9zNo ratings yet
- Pexman Et Al JoCN 2007Document13 pagesPexman Et Al JoCN 2007tkorotajdracaNo ratings yet
- Musical Form-Behaviour and NeuroDocument19 pagesMusical Form-Behaviour and NeuroMarcos EvereadyNo ratings yet
- Knoferle FinalDocument42 pagesKnoferle FinalJherleen Garcia CabacunganNo ratings yet
- 2011 CerebCortex 2011 Abrams 1507 18Document12 pages2011 CerebCortex 2011 Abrams 1507 18karmagonNo ratings yet
- Predicting Variation of Folk Songs A Corpus AnalysDocument12 pagesPredicting Variation of Folk Songs A Corpus AnalysRaven LaronNo ratings yet
- Neural Correlates of Accelerated Processing in Children Engaged in Music TrainingDocument14 pagesNeural Correlates of Accelerated Processing in Children Engaged in Music TrainingJosé GresalNo ratings yet
- TMP 304 CDocument27 pagesTMP 304 CFrontiersNo ratings yet
- Language, Brain and RepresentationDocument39 pagesLanguage, Brain and Representationsumitasthana2No ratings yet
- Bab 3Document5 pagesBab 3dickyginanjarNo ratings yet
- Slevc Okada 2015 PBR PostprintDocument31 pagesSlevc Okada 2015 PBR PostprintMiguel Iago CavalcantiNo ratings yet
- An Investigation Into The Temporal Dimension of The Mozart Effect - Evidence From The Attentional Blink TaskDocument12 pagesAn Investigation Into The Temporal Dimension of The Mozart Effect - Evidence From The Attentional Blink TaskmariottoalbertoNo ratings yet
- tmp30DE TMPDocument6 pagestmp30DE TMPFrontiersNo ratings yet
- 1 s2.0 S001094520870491X MainDocument9 pages1 s2.0 S001094520870491X MainAlberto EarnshawNo ratings yet
- Technology Predictions For 2025Document1 pageTechnology Predictions For 2025Sze Sze LiNo ratings yet
- Technology Predictions For 2025Document1 pageTechnology Predictions For 2025Sze Sze LiNo ratings yet
- Reading Chinese ScriptDocument1 pageReading Chinese ScriptSze Sze LiNo ratings yet
- Quantum Computing to Revolutionize Drug Development and Materials Design by 2025Document1 pageQuantum Computing to Revolutionize Drug Development and Materials Design by 2025Sze Sze LiNo ratings yet
- Modelling Chinese Character PronunciationDocument6 pagesModelling Chinese Character PronunciationSze Sze LiNo ratings yet
- Technology Predictions For 2025Document1 pageTechnology Predictions For 2025Sze Sze LiNo ratings yet
- Body and HealthDocument2 pagesBody and HealthSze Sze LiNo ratings yet
- Technology Predictions For 2025Document1 pageTechnology Predictions For 2025Sze Sze LiNo ratings yet
- Neurocognitive Approaches To Bilingualism - Asian LanguagesDocument3 pagesNeurocognitive Approaches To Bilingualism - Asian LanguagesSze Sze LiNo ratings yet
- Incremental Learning of Chinese OrthographyDocument18 pagesIncremental Learning of Chinese OrthographySze Sze LiNo ratings yet
- Writing Experiences On Holistic Processing in Chinese Character RecognitionDocument6 pagesWriting Experiences On Holistic Processing in Chinese Character RecognitionSze Sze LiNo ratings yet
- Moral Capitalism - Stakeholders Take Over From ShareholdersDocument5 pagesMoral Capitalism - Stakeholders Take Over From ShareholdersSze Sze LiNo ratings yet
- Neurocognitive Approaches To Bilingualism - Asian LanguagesDocument3 pagesNeurocognitive Approaches To Bilingualism - Asian LanguagesSze Sze LiNo ratings yet
- Incremental Learning of Chinese OrthographyDocument18 pagesIncremental Learning of Chinese OrthographySze Sze LiNo ratings yet
- Symbolic Representation For Reading in Asian and Alphabetic LanguagesDocument9 pagesSymbolic Representation For Reading in Asian and Alphabetic LanguagesSze Sze LiNo ratings yet
- How Mind Can Meet Brain in ReadingDocument14 pagesHow Mind Can Meet Brain in ReadingSze Sze LiNo ratings yet
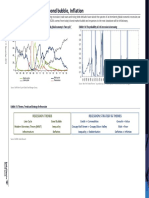
- Recession - Late Cycle, Bond Bubble, InflationDocument4 pagesRecession - Late Cycle, Bond Bubble, InflationSze Sze LiNo ratings yet
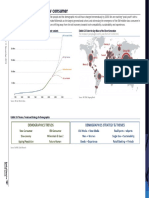
- Demographics - The New ConsumerDocument7 pagesDemographics - The New ConsumerSze Sze LiNo ratings yet
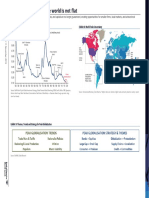
- Peak Globalization: Real Assets Ripe for Renewed InterestDocument4 pagesPeak Globalization: Real Assets Ripe for Renewed InterestSze Sze LiNo ratings yet
- Smart Everything - Ubiquitous Connectivity & Death of PrivacyDocument4 pagesSmart Everything - Ubiquitous Connectivity & Death of PrivacySze Sze LiNo ratings yet
- Splinternet - The Race For Technological Supremacy and Sovereign InternetsDocument6 pagesSplinternet - The Race For Technological Supremacy and Sovereign InternetsSze Sze LiNo ratings yet
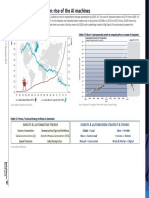
- Robots and Automation - Rise of The AI MachinesDocument4 pagesRobots and Automation - Rise of The AI MachinesSze Sze LiNo ratings yet
- Quantitative Failure - Post QE DecadeDocument4 pagesQuantitative Failure - Post QE DecadeSze Sze LiNo ratings yet
- New Paradigms of The 2020S: Exhibit 1: Bofaml Transforming World: 2010S vs. 2020SDocument1 pageNew Paradigms of The 2020S: Exhibit 1: Bofaml Transforming World: 2010S vs. 2020SSze Sze LiNo ratings yet
- Dyslexia HeterogeneityDocument17 pagesDyslexia HeterogeneitySze Sze LiNo ratings yet
- Araújo Et Al (2013) Phonological ProcessDocument16 pagesAraújo Et Al (2013) Phonological ProcessCarolinaNo ratings yet
- Did You Know in The Next DecadeDocument1 pageDid You Know in The Next DecadeSze Sze LiNo ratings yet
- Climate Change - Make Earth Green AgainDocument4 pagesClimate Change - Make Earth Green AgainSze Sze LiNo ratings yet
- Lexical and Sublexical Orthographic ProcessingDocument12 pagesLexical and Sublexical Orthographic ProcessingSze Sze LiNo ratings yet
- Ai, Iot, Big Data & BlockchainDocument19 pagesAi, Iot, Big Data & BlockchainClick GroupNo ratings yet
- Role of Digital Infrastructure in The Post Pandemic World: Preparing For Civil Services Interview-The Right PerspectiveDocument40 pagesRole of Digital Infrastructure in The Post Pandemic World: Preparing For Civil Services Interview-The Right PerspectiveshekhardfcNo ratings yet
- Ullit Présentation GénéraleDocument34 pagesUllit Présentation Généralepvk.p53No ratings yet
- Distributed Tic-Tac-Toe: Requirements, Design & AnalysisDocument8 pagesDistributed Tic-Tac-Toe: Requirements, Design & AnalysisRitika AgnihotriNo ratings yet
- Prince George's County Afro-American Newspaper, January 22, 2011Document16 pagesPrince George's County Afro-American Newspaper, January 22, 2011The AFRO-American NewspapersNo ratings yet
- 2.3 Motion and SafetyDocument13 pages2.3 Motion and SafetyGeorge TongNo ratings yet
- шпоры стилDocument19 pagesшпоры стилYuliia VasylchenkoNo ratings yet
- Crown Platform Crown Battery Operated Pallet TruckDocument7 pagesCrown Platform Crown Battery Operated Pallet Truckgaurav champawatNo ratings yet
- Toshiba SMMS-7 VRF CatalogueDocument106 pagesToshiba SMMS-7 VRF CatalogueMyo Sein67% (3)
- SMD - Metal-Sheet Decking DesignDocument10 pagesSMD - Metal-Sheet Decking DesignAchilles YipNo ratings yet
- 2022 AMC Paper MP OMRDocument12 pages2022 AMC Paper MP OMRshilpa_aryan100% (1)
- Lisa Van Krieken CVDocument1 pageLisa Van Krieken CVapi-525455549No ratings yet
- Evaluation of Earthquake-Induced Cracking of Embankment DamsDocument21 pagesEvaluation of Earthquake-Induced Cracking of Embankment DamsMARCOS ABRAHAM ALEJANDRO BALDOCEDA HUAYASNo ratings yet
- Quality Assurance AnalystDocument2 pagesQuality Assurance AnalystMalikNo ratings yet
- Geotechnical Scope of Work For ERL Unit-2Document75 pagesGeotechnical Scope of Work For ERL Unit-2smjohirNo ratings yet
- Monika Soni PDFDocument16 pagesMonika Soni PDFdeepak guptaNo ratings yet
- Class NoteDocument33 pagesClass NoteAnshuman MukherjeeNo ratings yet
- Robinair Mod 10324Document16 pagesRobinair Mod 10324StoneAge1No ratings yet
- Unleashing Learner PowerDocument188 pagesUnleashing Learner PowerEka MeilinaNo ratings yet
- Chapter Arithmetic and Geometric Progressions 024005Document14 pagesChapter Arithmetic and Geometric Progressions 024005ravichandran_brNo ratings yet
- An Introduction To Closed Loop MarketingDocument40 pagesAn Introduction To Closed Loop MarketingGeorgiana VasilescuNo ratings yet
- X20 System-ENG - V3.50 PDFDocument3,410 pagesX20 System-ENG - V3.50 PDFAbdelrahmanJamal100% (1)
- Troubleshooting Edge Quality: Mild SteelDocument14 pagesTroubleshooting Edge Quality: Mild SteelAnonymous U6yVe8YYCNo ratings yet
- Apollo System Engineering Manual. Environmental Control SystemDocument137 pagesApollo System Engineering Manual. Environmental Control Systemjackie_fisher_email8329100% (2)
- Presentation MolesworthDocument13 pagesPresentation Molesworthapi-630754890No ratings yet
- B. Glo Bal Maxima/MinimaDocument15 pagesB. Glo Bal Maxima/MinimalucasNo ratings yet
- ICs Interface and Motor DriversDocument4 pagesICs Interface and Motor DriversJohn Joshua MontañezNo ratings yet
- Centrifugal PumpDocument44 pagesCentrifugal PumpAmishaan KharbandaNo ratings yet
- BS en 13035-9-2006 + A1-2010Document24 pagesBS en 13035-9-2006 + A1-2010DoicielNo ratings yet
- Risktec Drilling Soc 0.3 PDFDocument16 pagesRisktec Drilling Soc 0.3 PDF1sNo ratings yet