You might also like
- Cell CycleDocument3 pagesCell Cycleapi-502781581No ratings yet
- Cells: Nuclear Envelope Proteins Modulating The Heterochromatin Formation and Functions in Fission YeastDocument21 pagesCells: Nuclear Envelope Proteins Modulating The Heterochromatin Formation and Functions in Fission YeastLê Khánh ToànNo ratings yet
- The Nuclear LaminaDocument27 pagesThe Nuclear Lamina박현탁 / 학생 / 기계공학부No ratings yet
- Lecture 2016 - Cell NucleusDocument17 pagesLecture 2016 - Cell NucleusKhalil RehmanNo ratings yet
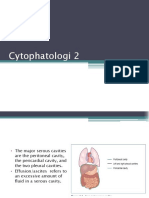
- CYTOPATHOLOGI 2 Pak DaniDocument40 pagesCYTOPATHOLOGI 2 Pak DaniYadi ApriyadiNo ratings yet
- Intermediate Filaments (Ifs)Document43 pagesIntermediate Filaments (Ifs)Baiq Kamelia Rahma Sari 019No ratings yet
- NucleusDocument9 pagesNucleussoumyadeepsahamondal20No ratings yet
- Cell Nucleus: Chromosome Dynamics in Nuclei of Living Cells: Daniele Zink and Thomas CremerDocument4 pagesCell Nucleus: Chromosome Dynamics in Nuclei of Living Cells: Daniele Zink and Thomas CremerThis is ScribeNo ratings yet
- Chapter 06Document39 pagesChapter 06NnleinomNo ratings yet
- Review Epigenetic Control of Nuclear Architecture: Cellular and Molecular Life SciencesDocument9 pagesReview Epigenetic Control of Nuclear Architecture: Cellular and Molecular Life SciencesPaige MunroeNo ratings yet
- A Hierarchical Combination of Factors Shapes The GDocument13 pagesA Hierarchical Combination of Factors Shapes The GMohan ParthasarathyNo ratings yet
- Pre-mRNA Splicing Life at The Centre of The Central DogmaDocument3 pagesPre-mRNA Splicing Life at The Centre of The Central DogmaQwyn Kym De GuzmanNo ratings yet
- Molecular Cell Biology 7Th Edition Full ChapterDocument41 pagesMolecular Cell Biology 7Th Edition Full Chapterjoseph.backman295100% (21)
- Ib Biology - Summary of "Draw, Label, Annotate" Assessment StatementsDocument3 pagesIb Biology - Summary of "Draw, Label, Annotate" Assessment StatementsJacky LeeNo ratings yet
- Artigo 1Document7 pagesArtigo 1GUSTAVO DOS SANTOS CARDOSONo ratings yet
- Iven So 2016Document9 pagesIven So 2016Ade Rizky FajrullohNo ratings yet
- Genome Organization and Function: A View From Yeast and ArabidopsisDocument13 pagesGenome Organization and Function: A View From Yeast and ArabidopsisJulienne InostrozaNo ratings yet
- Organization and Evolution of The Nuclear Genome: Princess Lorraine Garcia Bs-Biology IvDocument25 pagesOrganization and Evolution of The Nuclear Genome: Princess Lorraine Garcia Bs-Biology IvLouella ArtatesNo ratings yet
- 2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisDocument16 pages2019 - Off The Wall - The Rhyme and Reason of Neurospora Crassa Hyphal MorphogenesisskbucherNo ratings yet
- Replisome MechanicsDocument9 pagesReplisome MechanicsStevens GonzalezNo ratings yet
- Van Deventer Et Al - 2021Document11 pagesVan Deventer Et Al - 2021SusanaNo ratings yet
- Ribosome Structure and ShapeDocument7 pagesRibosome Structure and ShapeLutviana TiaswuniNo ratings yet
- Plant Genome OrganisationDocument16 pagesPlant Genome OrganisationVerônica D'AddazioNo ratings yet
- Development of The Cerebral Cortex: Forming The Cortical StructureDocument2 pagesDevelopment of The Cerebral Cortex: Forming The Cortical StructureMouhamad Al-kurdiNo ratings yet
- Rad9 Gene of Cinereus Encodes A Proline-Rich Protein RequiredDocument13 pagesRad9 Gene of Cinereus Encodes A Proline-Rich Protein RequiredAJMRNo ratings yet
- Pbifvjeoiif Ki Ojo LKJ JKNJ NJ Noi Jo Joi Jio Jiu JDocument3 pagesPbifvjeoiif Ki Ojo LKJ JKNJ NJ Noi Jo Joi Jio Jiu JLaia Baus RevueltaNo ratings yet
- Truskina 2018Document6 pagesTruskina 2018Supredo ManurungNo ratings yet
- High-Resolution Human Genome Structure by Single-Molecule AnalysisDocument6 pagesHigh-Resolution Human Genome Structure by Single-Molecule Analysisbrteag00No ratings yet
- Nuclear Architecture and The Structural Basis of Mitotic MemoryDocument23 pagesNuclear Architecture and The Structural Basis of Mitotic MemoryIrish Mae LunaNo ratings yet
- Pi Is 0006349518321933Document1 pagePi Is 0006349518321933ASP ValenciaNo ratings yet
- 2009, Mechanism Regulating S-Phase Progression in Mammalian CellsDocument15 pages2009, Mechanism Regulating S-Phase Progression in Mammalian CellsErnesto RojoNo ratings yet
- Fmicb 11 02115Document15 pagesFmicb 11 02115tejasNo ratings yet
- NIH Public Access: Author ManuscriptDocument25 pagesNIH Public Access: Author ManuscriptLokosooNo ratings yet
- EmergentDynamics PDFDocument10 pagesEmergentDynamics PDFBababa EkeNo ratings yet
- Lecture 16 3D OrganisationDocument27 pagesLecture 16 3D OrganisationNezrinNo ratings yet
- 15206Document190 pages15206cber_scribdNo ratings yet
- Replisome - WikipediaDocument17 pagesReplisome - WikipediaSibam KairiNo ratings yet
- Journal Pone 0028255Document13 pagesJournal Pone 0028255LeoNo ratings yet
- Cell Adhesion Molecules in Neural Stem Cell and Stem Cell-Based Therapy For Neural DisordersDocument32 pagesCell Adhesion Molecules in Neural Stem Cell and Stem Cell-Based Therapy For Neural DisordersQuang TranNo ratings yet
- Nucleus and ChromosomesDocument43 pagesNucleus and Chromosomesapi-19916399No ratings yet
- The Shoot Apical Meristem The Dynamics of A Stable StructureDocument11 pagesThe Shoot Apical Meristem The Dynamics of A Stable StructureMaribel PortelesNo ratings yet
- 2008 Synaptic Vesicle FusionDocument10 pages2008 Synaptic Vesicle FusionRidha Surya NugrahaNo ratings yet
- 2021-NP-What Shoots Can Teach About Theories of Plant FormDocument9 pages2021-NP-What Shoots Can Teach About Theories of Plant FormJay Prakash MauryaNo ratings yet
- 1a - Three Dimensional Genome Architecture Players and MechanismsDocument13 pages1a - Three Dimensional Genome Architecture Players and MechanismsStephaneNo ratings yet
- Kinetochore Assembly and Function Through The Cell Cycle: ReviewDocument15 pagesKinetochore Assembly and Function Through The Cell Cycle: ReviewLê Khánh ToànNo ratings yet
- MitosisDocument10 pagesMitosisbrxd69cmx7No ratings yet
- Mitosis: J. Richard McintoshDocument16 pagesMitosis: J. Richard McintoshLizbeth EspinosaNo ratings yet
- The Multifunctional Nucleolus: ReviewsDocument12 pagesThe Multifunctional Nucleolus: ReviewsleNo ratings yet
- Systematic Detection of Internal Symmetry in Proteins Using CE-SymmDocument14 pagesSystematic Detection of Internal Symmetry in Proteins Using CE-SymmelhadyegNo ratings yet
- Signorelli 2016Document10 pagesSignorelli 2016Wilson Agudelo CatañoNo ratings yet
- The Architecture of A Scrambled Genome Reveals Massive Levels of Genomic Rearrangement During DevelopmentDocument12 pagesThe Architecture of A Scrambled Genome Reveals Massive Levels of Genomic Rearrangement During DevelopmentAnonymous slVH85zYNo ratings yet
- Mitotic CheckpointsDocument32 pagesMitotic CheckpointsmikefirminoNo ratings yet
- 18 ChainDocument14 pages18 ChainMohit GargNo ratings yet
- Nuclear Pore ComplexesDocument9 pagesNuclear Pore ComplexesMNo ratings yet
- J Plrev 2011 02 005Document4 pagesJ Plrev 2011 02 005asdasddsaNo ratings yet
- Flujo de Información GenéticaDocument23 pagesFlujo de Información GenéticaKennyDeLosReyesNo ratings yet
- Annual Reviews - ReplicationDocument36 pagesAnnual Reviews - ReplicationfngermanoNo ratings yet
- Celladhesion Epithel KissADocument51 pagesCelladhesion Epithel KissAamdusuf33No ratings yet
- Lecture 3 TranscriptsDocument15 pagesLecture 3 TranscriptskittyngameNo ratings yet
- Emerin Modulates Spatial Organization of Chromosome Territories in Cells On Softer MatricesDocument26 pagesEmerin Modulates Spatial Organization of Chromosome Territories in Cells On Softer MatricesSubhadip KunduNo ratings yet
- 1 s2.0 S0960982209008094 MainDocument6 pages1 s2.0 S0960982209008094 MainLê Khánh ToànNo ratings yet
- Kinetochore Assembly and Function Through The Cell Cycle: ReviewDocument15 pagesKinetochore Assembly and Function Through The Cell Cycle: ReviewLê Khánh ToànNo ratings yet
- Alves Vietnamese Origins JVSDocument28 pagesAlves Vietnamese Origins JVSLê Khánh ToànNo ratings yet
- 2005 Ubx2 Links cdc48 To ERADDocument7 pages2005 Ubx2 Links cdc48 To ERADLê Khánh ToànNo ratings yet
- 2014 Nature PQC at INMDocument12 pages2014 Nature PQC at INMLê Khánh ToànNo ratings yet
- The Origin of Tones in Viet-Muong: To Cite This VersionDocument18 pagesThe Origin of Tones in Viet-Muong: To Cite This VersionLê Khánh ToànNo ratings yet
- 2020 QC Pathways of TA ProteinsDocument9 pages2020 QC Pathways of TA ProteinsLê Khánh ToànNo ratings yet
- A Cytosolic Degradation Pathway, Prerad, Monitors Pre-Inserted Secretory Pathway ProteinsDocument17 pagesA Cytosolic Degradation Pathway, Prerad, Monitors Pre-Inserted Secretory Pathway ProteinsLê Khánh ToànNo ratings yet
- 1998 Mata2 DegradationDocument11 pages1998 Mata2 DegradationLê Khánh ToànNo ratings yet
- 2011 TA Insertion Into ERDocument12 pages2011 TA Insertion Into ERLê Khánh ToànNo ratings yet
- Cytoplasmic Protein Quality Control Degradation Mediated by Parallel Actions of The E3 Ubiquitin Ligases Ubr1 and San1Document6 pagesCytoplasmic Protein Quality Control Degradation Mediated by Parallel Actions of The E3 Ubiquitin Ligases Ubr1 and San1Lê Khánh ToànNo ratings yet
- Cells 09 00518Document15 pagesCells 09 00518Lê Khánh ToànNo ratings yet
- Nuclear Membrane Rupture and Its Consequences: Annual Review of Cell and Developmental BiologyDocument30 pagesNuclear Membrane Rupture and Its Consequences: Annual Review of Cell and Developmental BiologyLê Khánh ToànNo ratings yet
- Degradation-Mediated Protein Quality Control in The Nucleus: Saccharomyces Cerevisiae. It Is Defined by San1p, ADocument13 pagesDegradation-Mediated Protein Quality Control in The Nucleus: Saccharomyces Cerevisiae. It Is Defined by San1p, ALê Khánh ToànNo ratings yet
- Beyond Tethering and The Lem Domain: Mscellaneous Functions of The Inner Nuclear Membrane Lem2Document10 pagesBeyond Tethering and The Lem Domain: Mscellaneous Functions of The Inner Nuclear Membrane Lem2Lê Khánh ToànNo ratings yet
- The Cohesin Complex in Mammalian Meiosis: Genes To CellsDocument25 pagesThe Cohesin Complex in Mammalian Meiosis: Genes To CellsLê Khánh ToànNo ratings yet
- The Initials of The Eastern Han Period As Reflected in Phonological GlossesDocument42 pagesThe Initials of The Eastern Han Period As Reflected in Phonological GlossesLê Khánh ToànNo ratings yet
- Biochimica Et Biophysica Acta: Ingo Amm, Thomas Sommer, Dieter H. WolfDocument15 pagesBiochimica Et Biophysica Acta: Ingo Amm, Thomas Sommer, Dieter H. WolfLê Khánh ToànNo ratings yet
- Letter: Distinct Proteostasis Circuits Cooperate in Nuclear and Cytoplasmic Protein Quality ControlDocument21 pagesLetter: Distinct Proteostasis Circuits Cooperate in Nuclear and Cytoplasmic Protein Quality ControlLê Khánh ToànNo ratings yet
- Nuclear Envelope Attachment Is Not Necessary For Telomere Function in Fission YeastDocument6 pagesNuclear Envelope Attachment Is Not Necessary For Telomere Function in Fission YeastLê Khánh ToànNo ratings yet
- ER-associated Degradation: Protein Quality Control and BeyondDocument11 pagesER-associated Degradation: Protein Quality Control and BeyondLê Khánh ToànNo ratings yet
- Chika 09 JCBDocument15 pagesChika 09 JCBLê Khánh ToànNo ratings yet
- Kenneth Gregerson - A Study of Middle Vietnamese PhonologyDocument64 pagesKenneth Gregerson - A Study of Middle Vietnamese PhonologyQuang-DungNo ratings yet
- Prehistoric Peopling of SEADocument6 pagesPrehistoric Peopling of SEALê Khánh ToànNo ratings yet
- Re-Imagining "Annam": A New Analysis of Sino-Viet - Muong Linguistic ContactDocument22 pagesRe-Imagining "Annam": A New Analysis of Sino-Viet - Muong Linguistic ContactLê Khánh ToànNo ratings yet
- Cell Function StructureDocument3 pagesCell Function StructureVeden Dumogho100% (1)
- Meiosis: Comes From The Greek Meioun, Meaning "To Make Smaller,"Document25 pagesMeiosis: Comes From The Greek Meioun, Meaning "To Make Smaller,"potionyesterday100% (2)
- IB Biology Revision SpreadsheetDocument124 pagesIB Biology Revision SpreadsheetTanika SharmaNo ratings yet
- Test Bank For Human Heredity Principles and Issues 10Th Edition Michael Cummings 1133106870 9781133106876 Full Chapter PDFDocument31 pagesTest Bank For Human Heredity Principles and Issues 10Th Edition Michael Cummings 1133106870 9781133106876 Full Chapter PDFkelly.rhodes162100% (13)
- General Zoology Module 1 Lesson 1-2.5Document5 pagesGeneral Zoology Module 1 Lesson 1-2.5Paul PanizaNo ratings yet
- Structure of Chromosomes, Cell Cycle, Cell DivisionDocument35 pagesStructure of Chromosomes, Cell Cycle, Cell DivisionXyzNo ratings yet
- CELL CYCLE McqsDocument2 pagesCELL CYCLE McqsHamzullah KhanNo ratings yet
- 3-Rajesh-Meiosis Class On 12th Feb 2013Document35 pages3-Rajesh-Meiosis Class On 12th Feb 2013Kanishk SinghNo ratings yet
- July 8 CXC BiologyDocument66 pagesJuly 8 CXC BiologyShantay BurtonNo ratings yet
- IJSO Biology Module - 1Document137 pagesIJSO Biology Module - 1Ikhbaat Atiqur RehmanNo ratings yet
- Genetics Lecture 2 Chromosomal Basis of HeredityDocument67 pagesGenetics Lecture 2 Chromosomal Basis of HeredityKareen Joy ManglicmotNo ratings yet
- Biology Form 3 Reproduction in Plants and Animals - Knec Study Materials, RevisDocument25 pagesBiology Form 3 Reproduction in Plants and Animals - Knec Study Materials, Reviskiraguripeter1994No ratings yet
- 01-Cell DivisionDocument21 pages01-Cell DivisionSENGUPTA CHANDANNo ratings yet
- Biology The Essentials 2nd Edition Mariëlle Hoefnagels Test Bank DownloadDocument38 pagesBiology The Essentials 2nd Edition Mariëlle Hoefnagels Test Bank DownloadGraham Hissem100% (21)
- เลคเชอร์รวมพันธุศาสตร์Document16 pagesเลคเชอร์รวมพันธุศาสตร์ณัฐภัทร วิศิษฎ์วรณัฐNo ratings yet
- WS Bio 10 Final Test 1st Sem 2019-20Document21 pagesWS Bio 10 Final Test 1st Sem 2019-20Jason Timothy KoesmanggalaNo ratings yet
- Knowing The Foundation of LifeDocument47 pagesKnowing The Foundation of LifeAlkhair SangcopanNo ratings yet
- AIFSET B.SC FS Model Test PaperDocument16 pagesAIFSET B.SC FS Model Test Paperkabita basakNo ratings yet
- PDFDocument4 pagesPDFYordan Gomez Capellan0% (1)
- Mitosis and Meiosis: Lecture Presentations by Cindy Malone California State University, NorthridgeDocument67 pagesMitosis and Meiosis: Lecture Presentations by Cindy Malone California State University, NorthridgeRicky Justin NgoNo ratings yet
- Bu2-3 Revision NotesDocument26 pagesBu2-3 Revision Notesveronica burlacuNo ratings yet
- All HSC Biology NotesDocument27 pagesAll HSC Biology Notesarabellatav23No ratings yet
- Onion Root Mitosis LabDocument6 pagesOnion Root Mitosis Labapi-2469736100% (1)
- Cell Division LabDocument8 pagesCell Division Labtworedpartyhats67% (6)
- SPM Mid Year 2008 SBP Biology Paper 1Document25 pagesSPM Mid Year 2008 SBP Biology Paper 1ChinWynn.com100% (1)
- Cell Division, 5.2Document6 pagesCell Division, 5.2PK100% (1)
- Kips FLP 7 MdcatDocument15 pagesKips FLP 7 MdcatIsmail HamzaNo ratings yet
- Unit 8 Cause & Effect, Comparison & Contrast, and Adverb ClausesDocument14 pagesUnit 8 Cause & Effect, Comparison & Contrast, and Adverb ClausescrrrrNo ratings yet
- SBL100 Minor I Questions AnswersDocument25 pagesSBL100 Minor I Questions AnswersApoorva DevNo ratings yet