You might also like
- Synthetic Biology and Metabolic EngineeringDocument8 pagesSynthetic Biology and Metabolic EngineeringLALUKISNo ratings yet
- (Springer Series in Wood Science) W. E. Hillis (auth.), John W. Rowe (eds.)-Natural Products of Woody Plants_ Chemicals Extraneous to the Lignocellulosic Cell Wall-Springer-Verlag Berlin Heidelberg (1.pdfDocument1,274 pages(Springer Series in Wood Science) W. E. Hillis (auth.), John W. Rowe (eds.)-Natural Products of Woody Plants_ Chemicals Extraneous to the Lignocellulosic Cell Wall-Springer-Verlag Berlin Heidelberg (1.pdfArmandoNo ratings yet
- BIO 328 Full Semester PackageDocument25 pagesBIO 328 Full Semester PackageNerdy Notes Inc.No ratings yet
- The Ethics of Respect For Nature - Paul W. Taylor PDFDocument22 pagesThe Ethics of Respect For Nature - Paul W. Taylor PDFArtur P. CoelhoNo ratings yet
- CHE 46: Biochemical EngineeringDocument30 pagesCHE 46: Biochemical EngineeringJunn Edgar LibotNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument16 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsIqbal PrawiraNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsHéctor EnriquezNo ratings yet
- Controlling Microbial Co-Culture Based On Substrate Pulsing Can Lead To Stability Through Differential Fitness AdvantagesDocument27 pagesControlling Microbial Co-Culture Based On Substrate Pulsing Can Lead To Stability Through Differential Fitness AdvantagesHamid MatariNo ratings yet
- Metabolomics Basic Principles and StrategiesDocument16 pagesMetabolomics Basic Principles and StrategiesEmiliano TorresNo ratings yet
- Chapter One Biotechnology and Biochemical EngineeringDocument22 pagesChapter One Biotechnology and Biochemical EngineeringYonas MegersaNo ratings yet
- Che 511 Lecture Note 2023Document5 pagesChe 511 Lecture Note 2023Bright ChimezieNo ratings yet
- Fib Unit 1 NotesDocument23 pagesFib Unit 1 Notespuja DhamijaNo ratings yet
- 1 Bioprocess Engineering CH 1Document22 pages1 Bioprocess Engineering CH 1Fasil ManNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument25 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsSrini VasanNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument41 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsOMiD QahqaieNo ratings yet
- Optimal AllocationDocument12 pagesOptimal AllocationRichardJGibsonNo ratings yet
- Lecture 23-Cell Growth Kinetics in Batch CultureDocument23 pagesLecture 23-Cell Growth Kinetics in Batch CultureHemanth Peddavenkatappa GariNo ratings yet
- Valorizing Food Grade Side Streams Using MicroorganismsDocument4 pagesValorizing Food Grade Side Streams Using MicroorganismsCharlotte LacrôixNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument29 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For Scientistskhue tranNo ratings yet
- Introduction 2013 Synthetic-BiologyDocument6 pagesIntroduction 2013 Synthetic-Biologyneo_hitmanNo ratings yet
- Escherichia Coli As A Model Organism and Its Application in Biotechnology PDFDocument23 pagesEscherichia Coli As A Model Organism and Its Application in Biotechnology PDFPerla AlvaradoNo ratings yet
- Conservación de AlimentosDocument42 pagesConservación de AlimentosDavid UreñaNo ratings yet
- Bioinspiration and Biomimicry in Chemistry: Reverse-Engineering NatureFrom EverandBioinspiration and Biomimicry in Chemistry: Reverse-Engineering NatureNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsUnnati GargNo ratings yet
- On The Organization of A Drug Discovery PlatformDocument39 pagesOn The Organization of A Drug Discovery PlatformBadou NindoNo ratings yet
- Utilizing Genetically Engineered OrganismsDocument13 pagesUtilizing Genetically Engineered OrganismsDidem KaraNo ratings yet
- Microbial Growth KineticsDocument20 pagesMicrobial Growth Kineticssushovan karNo ratings yet
- Trends in Biotechnology Volume 21 Issue 9 2003 [Doi 10.1016_s0167-7799(03)00190-2] Michael K Dyck_ Dan Lacroix_ François Pothier_ Marc-André Sira -- Making Recombinant Proteins in Animals – DifferentDocument6 pagesTrends in Biotechnology Volume 21 Issue 9 2003 [Doi 10.1016_s0167-7799(03)00190-2] Michael K Dyck_ Dan Lacroix_ François Pothier_ Marc-André Sira -- Making Recombinant Proteins in Animals – Differentahkhan086No ratings yet
- Krautkramer Metaboliti PDFDocument18 pagesKrautkramer Metaboliti PDFenaNo ratings yet
- The Bioreactor A Powerful Tool For Large-Scale Culture of Animal CellsDocument8 pagesThe Bioreactor A Powerful Tool For Large-Scale Culture of Animal CellsADITI AWASTHINo ratings yet
- Kinetics of Substrate Utilization, Product Formation and Biomass Production in Cell CulturesDocument5 pagesKinetics of Substrate Utilization, Product Formation and Biomass Production in Cell CulturesfayeNo ratings yet
- UntitledDocument12 pagesUntitledRaghavi ChinnaduraiNo ratings yet
- Leveraging Integrated Model-Based Approaches To UnDocument27 pagesLeveraging Integrated Model-Based Approaches To UnDiana TchoumiNo ratings yet
- 04 Computer Applications in Pharmacy Full Unit IVDocument14 pages04 Computer Applications in Pharmacy Full Unit IVSinh LuyenNo ratings yet
- Phenomics Workshop ReportDocument42 pagesPhenomics Workshop ReportEdin HamzicNo ratings yet
- Research in Global Environment-Cell and Molecular Biology: Sarthak Khurana AO572818003Document13 pagesResearch in Global Environment-Cell and Molecular Biology: Sarthak Khurana AO572818003sarthakNo ratings yet
- Jitorres - Lectura 3 CH02 - ASM - Montville - 557054 - 1-9Document9 pagesJitorres - Lectura 3 CH02 - ASM - Montville - 557054 - 1-9jelver andres pulido morenoNo ratings yet
- CHE 0412 1 - Topic 4 1Document62 pagesCHE 0412 1 - Topic 4 1Jelo ArtozaNo ratings yet
- Bio Film: Sowmya Subramanian, Ryan C. Huiszoon, Sangwook Chu, William E. Bentley, Reza GhodssiDocument16 pagesBio Film: Sowmya Subramanian, Ryan C. Huiszoon, Sangwook Chu, William E. Bentley, Reza GhodssiOussama El BouadiNo ratings yet
- Cell Culture Media-EbookDocument21 pagesCell Culture Media-EbookbioNo ratings yet
- Continuous Culture in ChemostatDocument6 pagesContinuous Culture in ChemostatBlessy GabaynoNo ratings yet
- C4CS00351ADocument68 pagesC4CS00351AVenkata Suryanarayana GorleNo ratings yet
- Production of Useful Secondary Metabolites Through Regulation of Biosynthetic Pathway in Cell and Tissue Suspension Culture of Medicinal PlantsDocument14 pagesProduction of Useful Secondary Metabolites Through Regulation of Biosynthetic Pathway in Cell and Tissue Suspension Culture of Medicinal Plantskj185No ratings yet
- 10 BookChapterGMOintechopenDocument25 pages10 BookChapterGMOintechopenSkittlessmannNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument21 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For Scientistskhalid bouitiNo ratings yet
- Sustainable Agriculture: Advances in Plant Metabolome and MicrobiomeFrom EverandSustainable Agriculture: Advances in Plant Metabolome and MicrobiomeNo ratings yet
- Food Microbiology Thesis PDFDocument7 pagesFood Microbiology Thesis PDFWriteMyNursingPaperTopeka100% (2)
- Bacterial Growth, Culturability and ViabilityDocument19 pagesBacterial Growth, Culturability and ViabilityRaúl Ernesto Quispe CórdovaNo ratings yet
- Metabolome Analysis For Investigating Host-Gut Microbiota InteractionsDocument13 pagesMetabolome Analysis For Investigating Host-Gut Microbiota InteractionssiscupNo ratings yet
- Expression Systems and Species Used For Transgenic Animal BioreactorsDocument10 pagesExpression Systems and Species Used For Transgenic Animal BioreactorsZeeshan AnjumNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument18 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsmusicontheroaddNo ratings yet
- An Introduction To The Reference Module in Life Sciences: Animal Physiology and BiologyDocument13 pagesAn Introduction To The Reference Module in Life Sciences: Animal Physiology and Biologyzouhir karimNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument15 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsSaad MaqboolNo ratings yet
- Biotechnology Progress - 2008 - Mandenius - Bioprocess Optimization Using Design of Experiments MethodologyDocument13 pagesBiotechnology Progress - 2008 - Mandenius - Bioprocess Optimization Using Design of Experiments MethodologyZahi DrawNo ratings yet
- Cytochrome P450 Monooxygenases in Biotechnology and Synthetic BiologyDocument58 pagesCytochrome P450 Monooxygenases in Biotechnology and Synthetic BiologyJaymuel Marco ResquirNo ratings yet
- Cell GrowthDocument48 pagesCell GrowthAlfin MartaNo ratings yet
- Microorganisms 06 00056Document12 pagesMicroorganisms 06 00056Juan Jesús Cruz MaldonadoNo ratings yet
- 1 Introduction To Biotechnology: 1.1 The Biotechnological ProcessDocument12 pages1 Introduction To Biotechnology: 1.1 The Biotechnological ProcessIshanSaneNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument29 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsMamathaNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument39 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistssumanchyNo ratings yet
- Research Paper On Fermentation TechnologyDocument5 pagesResearch Paper On Fermentation Technologyafedyvlyj100% (1)
- Metabolic Profiling of CHO Cells During The Production of BiotherapeuticsDocument21 pagesMetabolic Profiling of CHO Cells During The Production of Biotherapeuticsip87JCNo ratings yet
- Types of Fermentations: I-Liquid FermentationDocument28 pagesTypes of Fermentations: I-Liquid Fermentationnareman hassanNo ratings yet
- Task 02 Id No:181-025-031 Differentiate Between Different Types of Foodborne Illness Foodborne IllnessDocument1 pageTask 02 Id No:181-025-031 Differentiate Between Different Types of Foodborne Illness Foodborne IllnessRazia UrmiNo ratings yet
- TASK-3: Name Host Types of Diarrhe A Toxin Producin G Symptom SDocument2 pagesTASK-3: Name Host Types of Diarrhe A Toxin Producin G Symptom SRazia UrmiNo ratings yet
- Food Mbio II AssighnmentDocument10 pagesFood Mbio II AssighnmentRazia UrmiNo ratings yet
- Presentation1 Mbio 313Document12 pagesPresentation1 Mbio 313Razia UrmiNo ratings yet
- Food Mbio II AssighnmentDocument10 pagesFood Mbio II AssighnmentRazia UrmiNo ratings yet
- Course Title: Food Microbiology II Course Code: MBIO-402Document19 pagesCourse Title: Food Microbiology II Course Code: MBIO-402Razia UrmiNo ratings yet
- Mbio313 Assignment 01 181-025-031Document7 pagesMbio313 Assignment 01 181-025-031Razia UrmiNo ratings yet
- MBIO 404 Assighnment 02 181-025-031Document12 pagesMBIO 404 Assighnment 02 181-025-031Razia UrmiNo ratings yet
- Mbio313 Assignment 2Document8 pagesMbio313 Assignment 2Razia UrmiNo ratings yet
- MBIO 404 Assighnment 1 181-025-031Document8 pagesMBIO 404 Assighnment 1 181-025-031Razia UrmiNo ratings yet
- Mbio313 Assignment 01 181-025-031Document7 pagesMbio313 Assignment 01 181-025-031Razia UrmiNo ratings yet
- A New Species of Riverine Crab of The Genus Sundathelphusa Bott, 1969 (Crustacea: Brachyura: Gecarcinucidae) From Northeastern Luzon, PhilippinesDocument10 pagesA New Species of Riverine Crab of The Genus Sundathelphusa Bott, 1969 (Crustacea: Brachyura: Gecarcinucidae) From Northeastern Luzon, PhilippinesNel YagosNo ratings yet
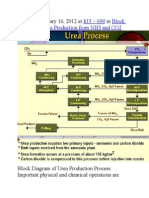
- Published January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2Document9 pagesPublished January 16, 2012 at In: 813 × 699 Block Diagram of Urea Production From NH3 and CO2himanshuchawla654No ratings yet
- The Impact of Your Internal Terrain On Your FasciaDocument16 pagesThe Impact of Your Internal Terrain On Your Fasciazzip1961100% (1)
- Temperament ThesisDocument4 pagesTemperament ThesisBuyCheapPaperOnlineSingapore100% (2)
- LeaP Science 10 Q4 W3 4Document5 pagesLeaP Science 10 Q4 W3 4D Garcia100% (2)
- Life Span: HumanDocument738 pagesLife Span: HumanWAJEEHA GOHARNo ratings yet
- Syllabus CompletionDocument1 pageSyllabus CompletiongopimicroNo ratings yet
- Sri Roth 2000Document11 pagesSri Roth 2000ottoojuniiorNo ratings yet
- PDFDocument570 pagesPDFEshaal FatimaNo ratings yet
- Pharyngeal ArchesDocument35 pagesPharyngeal ArchesconstantinilieNo ratings yet
- An Examination and Critique of Current Methods To Determine Exercise IntensityDocument28 pagesAn Examination and Critique of Current Methods To Determine Exercise IntensityPabloAñonNo ratings yet
- Clave Genero Dermanura (Otros) PDFDocument520 pagesClave Genero Dermanura (Otros) PDFJuan D Garcia Peluffo100% (1)
- Human Genetics Concepts and Applications 11Th Edition Ricki Lewis Test Bank Full Chapter PDFDocument36 pagesHuman Genetics Concepts and Applications 11Th Edition Ricki Lewis Test Bank Full Chapter PDFdavid.cordero230100% (14)
- The Tai Chi Code - Martial Arts Ebook by Chris DavisDocument190 pagesThe Tai Chi Code - Martial Arts Ebook by Chris DavisTomas De Wael100% (2)
- CLS Aipmt 15 16 XIII Zoo Study Package 2 Set 1 Chapter 3Document30 pagesCLS Aipmt 15 16 XIII Zoo Study Package 2 Set 1 Chapter 3Santu Karmakar100% (1)
- The Butterfly Circus Worksheet Activity - 68134Document3 pagesThe Butterfly Circus Worksheet Activity - 68134Hellguz ArtNo ratings yet
- Using STADEN For Sequence AssemblyDocument5 pagesUsing STADEN For Sequence AssemblyChristen Rune StensvoldNo ratings yet
- Taxonomy of Haematococcus PluvialisDocument2 pagesTaxonomy of Haematococcus PluvialisKomathi BalasupramaniamNo ratings yet
- Johnson Jerry Alan Chinese Medical Qigong Therapy Vol 5-141-160Document20 pagesJohnson Jerry Alan Chinese Medical Qigong Therapy Vol 5-141-160toanbauNo ratings yet
- Jagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Document305 pagesJagaroo & Santangelo - Neurophenotypes Advancing Psychiatry and Neuropsychology 2016Jaime Fernández-Aguirrebengoa100% (1)
- Jellyfish Research PaperDocument6 pagesJellyfish Research Papergzrvpcvnd100% (1)
- Tomasello, M. (2000) - Culture and Cognitive Development.Document4 pagesTomasello, M. (2000) - Culture and Cognitive Development.Roberto OcampoNo ratings yet
- Artikel SkripsiDocument12 pagesArtikel SkripsiSari RofiqohNo ratings yet
- Psychophysics As New Priority in Modern Science - Physical Basics of Informational InteractionDocument15 pagesPsychophysics As New Priority in Modern Science - Physical Basics of Informational InteractionBoris PetrovicNo ratings yet
- CROSS TREE - Sorte I Selekcije Voćaka I Vinove Loze Iz Severozapadne Srbije I Istočne Hrvatske PDFDocument153 pagesCROSS TREE - Sorte I Selekcije Voćaka I Vinove Loze Iz Severozapadne Srbije I Istočne Hrvatske PDFrraattaarrrraattaaNo ratings yet
- Indira Renganathan 2018 18Document1,992 pagesIndira Renganathan 2018 18shubham vedakNo ratings yet
- Prelim QuizDocument3 pagesPrelim QuizXyriel MacoyNo ratings yet



























![Trends in Biotechnology Volume 21 Issue 9 2003 [Doi 10.1016_s0167-7799(03)00190-2] Michael K Dyck_ Dan Lacroix_ François Pothier_ Marc-André Sira -- Making Recombinant Proteins in Animals – Different](https://imgv2-2-f.scribdassets.com/img/document/221613100/149x198/8025e9ddca/1399054344?v=1)