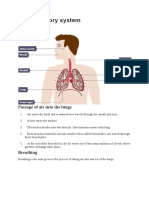
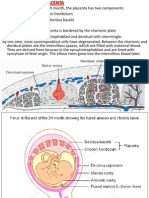
You might also like
- Treatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsFrom EverandTreatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsNo ratings yet
- 1 (Lung Development)Document12 pages1 (Lung Development)Dr.sonuNo ratings yet
- Neonatal Respiration PhysiologyDocument6 pagesNeonatal Respiration PhysiologyRaja Ahmad Rusdan MusyawirNo ratings yet
- Ultrasound in Obstet Gyne - 2002 - Laudy - The Fetal Lung 1 Developmental AspectsDocument7 pagesUltrasound in Obstet Gyne - 2002 - Laudy - The Fetal Lung 1 Developmental AspectsRafael JustinoNo ratings yet
- Pediatric and Neonatal Respiratory Care DevelopmentDocument205 pagesPediatric and Neonatal Respiratory Care DevelopmentAlexander Santiago ParelNo ratings yet
- Pulmonary EmbolismDocument21 pagesPulmonary EmbolismMadhu Bala100% (2)
- Development of the Respiratory System and Its AnomaliesDocument17 pagesDevelopment of the Respiratory System and Its AnomaliesmichaelqurtisNo ratings yet
- Lung Growth and Development: Suchita Joshi, Sailesh KotechaDocument6 pagesLung Growth and Development: Suchita Joshi, Sailesh KotechaAditya SanjayaNo ratings yet
- EMBRYOLOGY OF HEART AND LUNG DEVELOPMENTDocument7 pagesEMBRYOLOGY OF HEART AND LUNG DEVELOPMENTavinash dhameriyaNo ratings yet
- Learning Aims WWW - Embryology.ch/indexen - HTML: at The End of This Module You Should Be Able ToDocument13 pagesLearning Aims WWW - Embryology.ch/indexen - HTML: at The End of This Module You Should Be Able ToMuhammad Luqman HakimNo ratings yet
- All About RespiratoryDocument69 pagesAll About RespiratoryMarcellina Awing100% (1)
- Human Lung DevelopmentDocument20 pagesHuman Lung DevelopmentMila KarmilaNo ratings yet
- Bronchioles Then Divide Into Three TypesDocument3 pagesBronchioles Then Divide Into Three TypesJeremy EvansNo ratings yet
- Blood Vessels DevelopmentDocument4 pagesBlood Vessels Developmentمالك مناصرةNo ratings yet
- Neonatal Pulmonary PhysiologyDocument6 pagesNeonatal Pulmonary PhysiologyAnapaolaNo ratings yet
- The Respiratory System: Supplement To Text, Chapter 9Document77 pagesThe Respiratory System: Supplement To Text, Chapter 9Christina GonezNo ratings yet
- Respiratory EmbryologyDocument7 pagesRespiratory EmbryologySam JamNo ratings yet
- Development of Respiratory System: Dr. Saeed Vohra & Dr. Sanaa AlshaarawyDocument26 pagesDevelopment of Respiratory System: Dr. Saeed Vohra & Dr. Sanaa AlshaarawyChristineNo ratings yet
- Complications in The Transition From Fetal To Neonatal LifeDocument10 pagesComplications in The Transition From Fetal To Neonatal LifeCIRUABCNo ratings yet
- Development of The HeartDocument7 pagesDevelopment of The HeartJether Marc GardoseNo ratings yet
- Lecture 1 - Development of Respiratory SystemDocument27 pagesLecture 1 - Development of Respiratory SystemRachmad MusyaffaNo ratings yet
- Embryo Development Lungs+Body CavityDocument8 pagesEmbryo Development Lungs+Body CavityKingNo ratings yet
- Anatomy and Embryology of Umbilicus in Newborns: A Review and Clinical CorrelationsDocument7 pagesAnatomy and Embryology of Umbilicus in Newborns: A Review and Clinical Correlationsbayu pamungkasNo ratings yet
- Regulation of Placental VEGF and PlGF by OxygenDocument9 pagesRegulation of Placental VEGF and PlGF by OxygenFeri ZidaneNo ratings yet
- Neonatal Respiratory System ABBOTTDocument24 pagesNeonatal Respiratory System ABBOTTMario TGNo ratings yet
- Embryologi OF The Respiratory System: Arti Rosaria Dewi, DRDocument60 pagesEmbryologi OF The Respiratory System: Arti Rosaria Dewi, DRAsti NuriatiNo ratings yet
- Embryology Respiratory System: Formation of Lung BudsDocument1 pageEmbryology Respiratory System: Formation of Lung Budsrezairfan221No ratings yet
- Acute Respiratory Distres SyndromeDocument17 pagesAcute Respiratory Distres SyndromeLalrinchhani PautuNo ratings yet
- Chapter 53 - Exocrine Pancreas: Michael L. Steer M.DDocument70 pagesChapter 53 - Exocrine Pancreas: Michael L. Steer M.DpablenqueNo ratings yet
- Diagnostic Examination of Pulmonary TuberculosisDocument5 pagesDiagnostic Examination of Pulmonary TuberculosisMark BerbanoNo ratings yet
- 20 - Development of Respiratory SystemDocument33 pages20 - Development of Respiratory SystemDr.B.B.GosaiNo ratings yet
- Respiratory Disorders Case 1 AsthmaDocument21 pagesRespiratory Disorders Case 1 AsthmajoatasouzaNo ratings yet
- Respiratory Distress in The NewbornDocument146 pagesRespiratory Distress in The NewbornAmit RamrattanNo ratings yet
- Embryology Notes - emDocument18 pagesEmbryology Notes - emrigario80% (5)
- Development of Respiratory SystemDocument34 pagesDevelopment of Respiratory SystemNatalie HuiNo ratings yet
- Hernia DiafragmaDocument7 pagesHernia Diafragmasentaro2100% (1)
- PHS 402 Presentation v4Document24 pagesPHS 402 Presentation v4kinghenry882No ratings yet
- Development of The Respiratory SystemDocument8 pagesDevelopment of The Respiratory SystemDanielNo ratings yet
- Embryonic Development of The HeartDocument11 pagesEmbryonic Development of The HeartMatthew RyanNo ratings yet
- Update Fetal Circulation - MDM AmyDocument10 pagesUpdate Fetal Circulation - MDM AmyNana Yunus100% (1)
- Developmental Physiology - Lung Function During Growth and Development From Birth To Old Age by Calogero 2010Document15 pagesDevelopmental Physiology - Lung Function During Growth and Development From Birth To Old Age by Calogero 2010Fe TusNo ratings yet
- Embryology Course V - Body Cavities, Respiratory SystemDocument28 pagesEmbryology Course V - Body Cavities, Respiratory SystemRawa MuhsinNo ratings yet
- Blood VesselsDocument10 pagesBlood VesselsCaroleNo ratings yet
- Human Body SystemsDocument9 pagesHuman Body SystemsTiara Dwi PangestutiNo ratings yet
- 11th Lecture Amnion& Amniotic FluidDocument35 pages11th Lecture Amnion& Amniotic FluidHussein Al Saedi100% (1)
- Introduction To Physiological Changes in NEONATAL AND PEDIATRIC RESPICARE James Astrologo LamusaoDocument88 pagesIntroduction To Physiological Changes in NEONATAL AND PEDIATRIC RESPICARE James Astrologo LamusaoDanao, Aira A.No ratings yet
- Pleural EffusionDocument53 pagesPleural EffusionZhyraine Iraj D. CaluzaNo ratings yet
- Sumjv1i2p 174-179Document6 pagesSumjv1i2p 174-179Gregory JamesNo ratings yet
- MatthewVaracalloMD AnatomyThoraxBronchialDocument10 pagesMatthewVaracalloMD AnatomyThoraxBronchialLawson Mawulolo SetsofiaNo ratings yet
- Respiratory System EmbDocument29 pagesRespiratory System EmbDanish GujjarNo ratings yet
- Pathophysiology of RDSDocument5 pagesPathophysiology of RDSsandyNo ratings yet
- Corticosteroid in Lung MaturationDocument12 pagesCorticosteroid in Lung Maturationtommy japolaNo ratings yet
- Development of The Body Cavities: EmbryologyDocument9 pagesDevelopment of The Body Cavities: EmbryologyRania FadillaNo ratings yet
- Kaufmann-2004-Aspects of Human Fetoplacental V PDFDocument13 pagesKaufmann-2004-Aspects of Human Fetoplacental V PDFNguyễn Cao VânNo ratings yet
- Ribs Vertebral Column Sternum Abdominal Cavity Diaphragm Lungs Heart Blood Arteries VeinsDocument11 pagesRibs Vertebral Column Sternum Abdominal Cavity Diaphragm Lungs Heart Blood Arteries VeinsHamda HassanNo ratings yet
- First Page PDFDocument1 pageFirst Page PDFIndira Damar PangestuNo ratings yet
- 12 Lung DevelopmentDocument36 pages12 Lung Developmentf3er3No ratings yet
- Case VirchowsDocument14 pagesCase VirchowsShanti IntansariNo ratings yet
- Fetalphysiologyandthe Transitiontoextrauterine Life: Sarah U. Morton,, Dara BrodskyDocument13 pagesFetalphysiologyandthe Transitiontoextrauterine Life: Sarah U. Morton,, Dara BrodskyMarcela RodriguezNo ratings yet
- Termorregulacion NeonatosDocument5 pagesTermorregulacion NeonatosMarcela RodriguezNo ratings yet
- Moore Et Al-2016-Cochrane Database of Systematic ReviewsDocument125 pagesMoore Et Al-2016-Cochrane Database of Systematic ReviewsMarcela RodriguezNo ratings yet
- Contacto Piel A Piel en HipotermiaDocument5 pagesContacto Piel A Piel en HipotermiaMarcela RodriguezNo ratings yet
- Estabilidad Termica Del Prematuro en UcinDocument5 pagesEstabilidad Termica Del Prematuro en UcinMarcela RodriguezNo ratings yet
- Lifengetal FeasibilityAcceptabilityandEffectsofaFootSelf-CareEducationalIntervention2013Document9 pagesLifengetal FeasibilityAcceptabilityandEffectsofaFootSelf-CareEducationalIntervention2013Marcela RodriguezNo ratings yet
- ERC 2015. Section 6. Paediatric Life Supportian PDFDocument26 pagesERC 2015. Section 6. Paediatric Life Supportian PDFSergioFernandezUreñaNo ratings yet
- Effects of A Weighted Maternally-ScentedDocument8 pagesEffects of A Weighted Maternally-ScentedMarcela RodriguezNo ratings yet
- ReferenciasDocument1 pageReferenciasMarcela RodriguezNo ratings yet
- A Phase 2 Quasi-Experimental Trial Evaluating The Feasibility, AcceptabilityDocument9 pagesA Phase 2 Quasi-Experimental Trial Evaluating The Feasibility, AcceptabilityMarcela RodriguezNo ratings yet
- A Phase 2 Quasi-Experimental Trial Evaluating The Feasibility, AcceptabilityDocument9 pagesA Phase 2 Quasi-Experimental Trial Evaluating The Feasibility, AcceptabilityMarcela RodriguezNo ratings yet
- Summary of Benefits: Northeast RegionDocument16 pagesSummary of Benefits: Northeast RegionDavid NoreñaNo ratings yet
- Core 13 UcspDocument3 pagesCore 13 UcspJanice DomingoNo ratings yet
- Schermafbeelding 2021-09-18 Om 15.57.50Document386 pagesSchermafbeelding 2021-09-18 Om 15.57.50Esoteric Sensations100% (1)
- Chairs and ArchitectsDocument26 pagesChairs and ArchitectsStefan PalaghiaNo ratings yet
- Acute cholecystitis pathogenesis, diagnosis and clinical featuresDocument38 pagesAcute cholecystitis pathogenesis, diagnosis and clinical featuresAdrian BăloiNo ratings yet
- Grade 10 Sasmo: secθ + tanθ = q, secθ q. q q q qDocument3 pagesGrade 10 Sasmo: secθ + tanθ = q, secθ q. q q q qAje GhazaNo ratings yet
- Hybrid - 01Document209 pagesHybrid - 01Santos ArwinNo ratings yet
- Quail Eggs IncubatorDocument5 pagesQuail Eggs IncubatorMada Sanjaya WsNo ratings yet
- 2 Saliva PhysiologyDocument20 pages2 Saliva PhysiologyvelangniNo ratings yet
- Eating Disorders Reading AssignmentDocument2 pagesEating Disorders Reading AssignmentNasratullah sahebzadaNo ratings yet
- Feasibility Studies: Presenter: Imee G. ApostolDocument35 pagesFeasibility Studies: Presenter: Imee G. ApostolKaye Anne TorresNo ratings yet
- Punjab govt grants EOL to doctors for postgraduate trainingDocument2 pagesPunjab govt grants EOL to doctors for postgraduate trainingMasroor HassanNo ratings yet
- AI in Agile Project Management - v0.1Document48 pagesAI in Agile Project Management - v0.1Pedro LopesNo ratings yet
- Din Ling, 'Miss Sophia's Diary'Document2 pagesDin Ling, 'Miss Sophia's Diary'StudenteNo ratings yet
- Internal and External Institutions and Influences of Corporate GovernanceDocument39 pagesInternal and External Institutions and Influences of Corporate GovernanceLovely PasatiempoNo ratings yet
- Just Say NO To Baby EinsteinDocument5 pagesJust Say NO To Baby Einsteinsmartypinto0% (1)
- The Six Phase Meltdown CycleDocument28 pagesThe Six Phase Meltdown CycleIrene C. CruzNo ratings yet
- Lol Wellness Principles For Dynamic LivingDocument171 pagesLol Wellness Principles For Dynamic LivingTumwine Kahweza ProsperNo ratings yet
- Puppets in GreeceDocument6 pagesPuppets in GreeceStathis Markopoulos100% (1)
- An Instrument of Revival BABALOLADocument46 pagesAn Instrument of Revival BABALOLAGoriola David Gbolahan83% (6)
- ALD PatriciaLockwood TheCommunalMindDocument16 pagesALD PatriciaLockwood TheCommunalMindtillhopstockNo ratings yet
- Poka YokeDocument25 pagesPoka YokepatilpradnyaNo ratings yet
- Truth V.S. OpinionDocument19 pagesTruth V.S. OpinionBek Ah100% (2)
- 3 Week Lit 121Document48 pages3 Week Lit 121Cassandra Dianne Ferolino MacadoNo ratings yet
- A Review of Research On Teacher Beliefs and PracticesDocument20 pagesA Review of Research On Teacher Beliefs and Practiceschikondi sepulaNo ratings yet
- PROM PPT UpdatedDocument26 pagesPROM PPT Updatedsaurabh chauhanNo ratings yet
- Types of Communication Networks - StudytonightDocument10 pagesTypes of Communication Networks - StudytonightHAMEED ULLAHNo ratings yet
- English - Exam ECO - 2Document6 pagesEnglish - Exam ECO - 2yogie yohansyahNo ratings yet
- FH 2016 10 Ir PDFDocument59 pagesFH 2016 10 Ir PDFdanialNo ratings yet
- Journal of Cleaner Production: Fabíola Negreiros de Oliveira, Adriana Leiras, Paula CerynoDocument15 pagesJournal of Cleaner Production: Fabíola Negreiros de Oliveira, Adriana Leiras, Paula CerynohbNo ratings yet