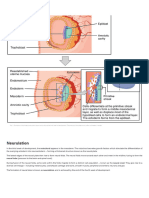
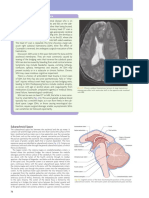
You might also like
- Chapter 50 Anatomy and Physiology of The NSDocument13 pagesChapter 50 Anatomy and Physiology of The NSana salamancaNo ratings yet
- Nervioso First AidDocument62 pagesNervioso First Aidadrenalina1238No ratings yet
- Brachial PlexusDocument5 pagesBrachial PlexusRajveerNo ratings yet
- Patterning The Developing DiencephalonDocument10 pagesPatterning The Developing DiencephalonAndres MoraNo ratings yet
- 1 OrientationDocument4 pages1 OrientationLeilani NajeraNo ratings yet
- Brain & Its Surrounding Structures: Anatomi Blok 1.5Document6 pagesBrain & Its Surrounding Structures: Anatomi Blok 1.5RizkiWikantyasningNo ratings yet
- Brain Encephalon: - Brain: Embrionally Comes From Ectoderm PartDocument72 pagesBrain Encephalon: - Brain: Embrionally Comes From Ectoderm PartHana AdivaNo ratings yet
- Salivary Glands: Fig. 11.1 Histologic Structure of The Normal Salivary GlandDocument35 pagesSalivary Glands: Fig. 11.1 Histologic Structure of The Normal Salivary GlandwillyoueverlovemenkNo ratings yet
- La Vía Piramidal: Nuevas Trayectorias: M.L. Cuadrado, J.A. Arias, M.A. Palomar, R. LinaresDocument8 pagesLa Vía Piramidal: Nuevas Trayectorias: M.L. Cuadrado, J.A. Arias, M.A. Palomar, R. Linarescl.lexisalazar00No ratings yet
- Cranial Nerves V, Ix, X, XiDocument89 pagesCranial Nerves V, Ix, X, XiUloko Christopher100% (1)
- Fig. 17.3B: RhombencephalonDocument2 pagesFig. 17.3B: RhombencephalonJSNo ratings yet
- Hubs2103 - Neural and Visceral AnatomyDocument153 pagesHubs2103 - Neural and Visceral AnatomyLuke ThomsonNo ratings yet
- Development of The Central Nervous System - SpinaDocument4 pagesDevelopment of The Central Nervous System - SpinaSoumya RathNo ratings yet
- EmbryologyDocument92 pagesEmbryologyLoveena SteadmanNo ratings yet
- Cerebrospinal Fluid RhinorrheaDocument4 pagesCerebrospinal Fluid RhinorrheaUlises VillanuevaNo ratings yet
- Clinical Neuroanatomical TechniquesDocument6 pagesClinical Neuroanatomical TechniquesMWNo ratings yet
- Nervo FacialeDocument15 pagesNervo FacialeAndrea PescosolidoNo ratings yet
- Mini AtlasDocument28 pagesMini AtlasAlex WoodwardNo ratings yet
- NBSS CASE 6 - Cerebral Palsy and Mental RetardationDocument28 pagesNBSS CASE 6 - Cerebral Palsy and Mental RetardationbungadielaNo ratings yet
- Neurology and Special Senses: High-Yield SystemsDocument72 pagesNeurology and Special Senses: High-Yield SystemsMahmoud Abu MayalehNo ratings yet
- Language and The CerebellumDocument22 pagesLanguage and The CerebellumDiane MxNo ratings yet
- Chapter 9 - The Occipital LobeDocument35 pagesChapter 9 - The Occipital LobeMorteza Mazloom Farsi BafNo ratings yet
- Anatomi Saraf New - 1Document61 pagesAnatomi Saraf New - 1Ahmad Ridwan JNo ratings yet
- Transient Structures, Subsequently Developing Into The Diverse Components of The Face and NeckDocument15 pagesTransient Structures, Subsequently Developing Into The Diverse Components of The Face and NecknohacksNo ratings yet
- Yanuzzi Retina Atlas: 1. Normal RetinaDocument6 pagesYanuzzi Retina Atlas: 1. Normal RetinaJorge Rendón F.No ratings yet
- Week 03a Insect HeadDocument5 pagesWeek 03a Insect HeadVanshika ChaudharyNo ratings yet
- Differentiation of Brain Vesicles Diencephalon PDFDocument27 pagesDifferentiation of Brain Vesicles Diencephalon PDFCony FigueroaNo ratings yet
- DEVELOPMENT NERVOUS SYSTEM Brain Face PDFDocument15 pagesDEVELOPMENT NERVOUS SYSTEM Brain Face PDFChidera EmmanuelNo ratings yet
- Anatomi Neuro PDFDocument106 pagesAnatomi Neuro PDFsciencewarriorNo ratings yet
- Exercise 4 Frog Embryo 4mm 7mm 10mmDocument23 pagesExercise 4 Frog Embryo 4mm 7mm 10mmrexartooz95% (19)
- Anatomi Sistem SarafDocument61 pagesAnatomi Sistem SarafMashuri MashuriNo ratings yet
- 72 HrsDocument35 pages72 HrsChristalie Bea FernandezNo ratings yet
- Bells PalsyDocument6 pagesBells PalsyPhub TsheringNo ratings yet
- Chapter 8Document4 pagesChapter 8Mia Kristhyn Calinawagan SabanalNo ratings yet
- Ebbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesDocument39 pagesEbbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesLivia PereiraNo ratings yet
- Lateral Cep Halo CeleDocument4 pagesLateral Cep Halo CeleAde Cahyo IslamiNo ratings yet
- Describe The Different Types of Ganglia in The Head and NeckDocument2 pagesDescribe The Different Types of Ganglia in The Head and Neckmp658t952dNo ratings yet
- Cortex and Supranuclear PathwayDocument14 pagesCortex and Supranuclear PathwayMohammed RahmathNo ratings yet
- Nerve Supply of Head and NeckDocument74 pagesNerve Supply of Head and NeckDr Komal GhiyaNo ratings yet
- Embryology Summary of Development Part 1 - 2019 - CNS, Axial SK, SK Muscle, SkinDocument9 pagesEmbryology Summary of Development Part 1 - 2019 - CNS, Axial SK, SK Muscle, Skindr.mumtaz09No ratings yet
- Ward Roger2001Document6 pagesWard Roger2001AntonyNo ratings yet
- Review Anatomi Fisiologi Sistem PersarafanDocument78 pagesReview Anatomi Fisiologi Sistem PersarafanNisa FathonahNo ratings yet
- Genesis Avian Cells: Classic Embryonic Induction: The of Neural Crest ADocument6 pagesGenesis Avian Cells: Classic Embryonic Induction: The of Neural Crest ARANJAN KUMARNo ratings yet
- Spinal Cord NS2.1 Final ArtDocument11 pagesSpinal Cord NS2.1 Final Artdedhomosapien2No ratings yet
- 33 Hour Chick Reviewer PDFDocument5 pages33 Hour Chick Reviewer PDFJedd VirgoNo ratings yet
- Glossopharyngeal Nerve: © L. Wilson-PauwelsDocument18 pagesGlossopharyngeal Nerve: © L. Wilson-PauwelsNorawaylandNo ratings yet
- Trigeminal Nerve and Its Applied AspectsDocument198 pagesTrigeminal Nerve and Its Applied AspectsArun MamachanNo ratings yet
- MeningesDocument43 pagesMeningesRehab NaeemNo ratings yet
- Trigeminal Nerve Seminar FINALDocument54 pagesTrigeminal Nerve Seminar FINALAnji Satsangi100% (1)
- Lesson 1 EyesDocument8 pagesLesson 1 EyesMARY JANE ANGELICA SEVANo ratings yet
- Baskin 2003Document5 pagesBaskin 2003Maria Luiza OliveiraNo ratings yet
- 33 HR Chick EmbryoDocument12 pages33 HR Chick EmbryoErvin AblazaNo ratings yet
- The Pyramidal Tract: New Pathways: Revista de Neurologia January 2001Document9 pagesThe Pyramidal Tract: New Pathways: Revista de Neurologia January 2001javier solisNo ratings yet
- Anatomi Asyik FkuiiDocument14 pagesAnatomi Asyik Fkuiibima_uiiNo ratings yet
- Cranial CavityDocument94 pagesCranial CavityShashank Tripathi100% (1)
- Growth and Development of Cranial Base: SeminarDocument34 pagesGrowth and Development of Cranial Base: SeminarMNSNo ratings yet
- Exp-01 A-14 Ashiya ThakurDocument15 pagesExp-01 A-14 Ashiya ThakurAshiya ThakurNo ratings yet
- Major Divisions of The Nervous SystemDocument38 pagesMajor Divisions of The Nervous SystemRiyadh AlhaddadNo ratings yet
- CNS Study GuideDocument6 pagesCNS Study GuideNasibah BaruangNo ratings yet
- Case 3 S H: Ubdural AematomaDocument9 pagesCase 3 S H: Ubdural AematomaAndreea LăzăroiuNo ratings yet
- References: Section I / GeneralDocument10 pagesReferences: Section I / GeneralAndreea LăzăroiuNo ratings yet
- Acknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsDocument7 pagesAcknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsAndreea LăzăroiuNo ratings yet
- Otic Ganglion: Schema: Plate 145Document8 pagesOtic Ganglion: Schema: Plate 145Andreea LăzăroiuNo ratings yet
- Case 1 A S: Queductal TenosisDocument11 pagesCase 1 A S: Queductal TenosisAndreea LăzăroiuNo ratings yet
- Medial Wall of Nasal Cavity (Nasal Septum) : Plate 15Document11 pagesMedial Wall of Nasal Cavity (Nasal Septum) : Plate 15Andreea LăzăroiuNo ratings yet
- The Atomic Divergence: Nuclear Energy Seems To Offer Some AdvantagesDocument3 pagesThe Atomic Divergence: Nuclear Energy Seems To Offer Some AdvantagesAndreea LăzăroiuNo ratings yet
- Exam Practice 1: Reading: PHOTOCOP' Sa 0 Education Limited 2008Document6 pagesExam Practice 1: Reading: PHOTOCOP' Sa 0 Education Limited 2008Andreea LăzăroiuNo ratings yet
- Exam Practice: 1, TRB p.178Document1 pageExam Practice: 1, TRB p.178Andreea LăzăroiuNo ratings yet
- Cerebellum (Section 3, Chapter 5) Neuroscience Online - An Electronic Textbook For The Neurosciences - Department of Neurobiology and Anatomy - The University of Texas Medical School at HoustonDocument9 pagesCerebellum (Section 3, Chapter 5) Neuroscience Online - An Electronic Textbook For The Neurosciences - Department of Neurobiology and Anatomy - The University of Texas Medical School at HoustonNaveen KumarNo ratings yet
- Naheed Ali, Moshe Lewis-Understanding Pain - An Introduction For Patients and Caregivers-Rowman & Littlefield Publishers (2015)Document415 pagesNaheed Ali, Moshe Lewis-Understanding Pain - An Introduction For Patients and Caregivers-Rowman & Littlefield Publishers (2015)MaryNo ratings yet
- Overview: Functions of The CerebellumDocument17 pagesOverview: Functions of The CerebellumMohd Fitri ShahruddinNo ratings yet
- 1971 Albus - A Theory of Cerebellar FunctionDocument37 pages1971 Albus - A Theory of Cerebellar FunctionmarkmcwilliamsNo ratings yet
- Childhood Neuronal Ceroid-Lipofuscinoses Argentina: 1995 Wiley-Liss, IncDocument6 pagesChildhood Neuronal Ceroid-Lipofuscinoses Argentina: 1995 Wiley-Liss, IncMaxime PorcoNo ratings yet
- Understanding PainDocument415 pagesUnderstanding PainShreyasi PatankarNo ratings yet
- Cell Death in Weaver Mouse Cerebellum: Amy B Harkins and Aaron P FoxDocument6 pagesCell Death in Weaver Mouse Cerebellum: Amy B Harkins and Aaron P FoxdupuytrenNo ratings yet
- The Giant First YearDocument1,234 pagesThe Giant First YearEthan DyerNo ratings yet
- Neurogenesis en El Cerebro AdultoDocument8 pagesNeurogenesis en El Cerebro AdultoNicolle Constanza Reiher MaierNo ratings yet
- Neural Plasticity in The Ageing BrainDocument11 pagesNeural Plasticity in The Ageing BrainGuillermo AranedaNo ratings yet
- Joseph D. Robinson - Mechanisms of Synaptic Transmission - Bridging The Gaps (1890-1990) (2001)Document468 pagesJoseph D. Robinson - Mechanisms of Synaptic Transmission - Bridging The Gaps (1890-1990) (2001)Pablo EspinosaNo ratings yet
- Physiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamDocument61 pagesPhysiology of Cerebellum Lec Foe 2nd Year by DR Sadia Uploaded by ZaighamZaigham HammadNo ratings yet
- Nervesandpsychiatry PDFDocument822 pagesNervesandpsychiatry PDFGruff Gruffer100% (1)
- David Attwell - Pericytes in Neurovascular Coupling in Health and DiseaseDocument39 pagesDavid Attwell - Pericytes in Neurovascular Coupling in Health and DiseaseFondation Singer-PolignacNo ratings yet
- Cephalopod Neural NetworksDocument12 pagesCephalopod Neural NetworksSabrina Indri WardaniNo ratings yet
- Neurodegenerative Cerebellar AtaxiaDocument26 pagesNeurodegenerative Cerebellar AtaxiaМилица МилошевићNo ratings yet
- Clark Et Al 2010Document7 pagesClark Et Al 2010fnmendoncaNo ratings yet
- Craig and Banker 1994Document44 pagesCraig and Banker 1994Kanwal RashidNo ratings yet
- Special HistologyDocument64 pagesSpecial HistologyElijah KamaniNo ratings yet
- Embriology Hewan - Michael - J. - F. - Barresi - Scott - F. - Gilbert - Developm (B-Ok - Xyz) (2) - 467-490Document24 pagesEmbriology Hewan - Michael - J. - F. - Barresi - Scott - F. - Gilbert - Developm (B-Ok - Xyz) (2) - 467-490Afriani AfidahNo ratings yet
- Lecture On PHYSIOLOGY of Cerebellum by Dr. RoomiDocument42 pagesLecture On PHYSIOLOGY of Cerebellum by Dr. RoomiMudassar Roomi100% (2)
- CEREBELLUM - Class 2Document14 pagesCEREBELLUM - Class 228g8jcfcqfNo ratings yet
- The Hippocampus BookDocument853 pagesThe Hippocampus BookLucia Lopez100% (3)
- Histology of Spinal Cord and Cerebellum by Dr. ROOMIDocument21 pagesHistology of Spinal Cord and Cerebellum by Dr. ROOMIMudassar Roomi100% (1)
- The Underpinnings of Cerebellar AtaxiasDocument16 pagesThe Underpinnings of Cerebellar AtaxiasManuek GarciaNo ratings yet
- 10 CerebellumDocument26 pages10 CerebellumDarpan KumariNo ratings yet
- INICET November 2021Document244 pagesINICET November 2021sonali bhakhoreeNo ratings yet
- Clinical: Dialogu S eDocument144 pagesClinical: Dialogu S eDiego A. Cuadros Torres100% (3)
- 2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusDocument8 pages2008 - Ehninger, Kempermann - Neurogenesis in The Adult HippocampusNityananda PortelladaNo ratings yet
- Table 1. General Somatic Sensory: Tracts Stimulus Function N1 N2 Decussation N3 N4 Additional InfoDocument8 pagesTable 1. General Somatic Sensory: Tracts Stimulus Function N1 N2 Decussation N3 N4 Additional InfoNicole Villaflor FabicoNo ratings yet