You might also like
- 10.1016@S0079 61230661011 XDocument13 pages10.1016@S0079 61230661011 XCoNo ratings yet
- Neuronal Nitric Oxide Synthase Expressin PDFDocument16 pagesNeuronal Nitric Oxide Synthase Expressin PDFShaneNo ratings yet
- Francaet Al BrainRes PDFDocument13 pagesFrancaet Al BrainRes PDFPrema Bhakti DasNo ratings yet
- Schmidt 1996Document8 pagesSchmidt 1996M4shroomNo ratings yet
- Arndt 1996Document27 pagesArndt 1996M4shroomNo ratings yet
- Las Espinas DendríticasDocument43 pagesLas Espinas DendríticasElíasNo ratings yet
- TRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisDocument10 pagesTRKB Downregulation Is Required For Dendrite Retraction in Developing Neurons of Chicken Nucleus MagnocellularisFrontiersNo ratings yet
- 10 1002@sici1096-986119960812372@188@@aid-Cne73 0 Co2-IDocument23 pages10 1002@sici1096-986119960812372@188@@aid-Cne73 0 Co2-IM4shroomNo ratings yet
- The Anatomical Record - 2013 - Hern Ndez Morato - Somatotopy of The Neurons Innervating The Cricothyroid PosteriorDocument10 pagesThe Anatomical Record - 2013 - Hern Ndez Morato - Somatotopy of The Neurons Innervating The Cricothyroid PosteriormanuelNo ratings yet
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDocument34 pagesThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337No ratings yet
- Neural Crest: Contributions To The Development of The Vertebrate HeadDocument10 pagesNeural Crest: Contributions To The Development of The Vertebrate HeadKanish AggarwalNo ratings yet
- Carr Central ProjectionsDocument13 pagesCarr Central ProjectionsMengda ZhangNo ratings yet
- Jurnal PendidikanDocument8 pagesJurnal PendidikanQothrun Nada FYNo ratings yet
- Tekhnik Pewarnaan Neuron Dan Neuroglia Pada Sistem Saraf PusatDocument8 pagesTekhnik Pewarnaan Neuron Dan Neuroglia Pada Sistem Saraf PusatNayna AfridaNo ratings yet
- Arroyo_J_Neurosci_1999Document13 pagesArroyo_J_Neurosci_1999Constanza FuentesNo ratings yet
- Module 2 Post Task 1Document1 pageModule 2 Post Task 1KAIRA FLORENZIA D. BANDONGNo ratings yet
- Interneurons in The Developing Human Neocortex: Nada Zecevic, Frances Hu, Igor JakovcevskiDocument16 pagesInterneurons in The Developing Human Neocortex: Nada Zecevic, Frances Hu, Igor JakovcevskiFrontiersNo ratings yet
- TMP 3151Document12 pagesTMP 3151FrontiersNo ratings yet
- The Dorsal Raphe Nucleus and Serotonin - Implications For Neuroplasticity Linked To Major Depression and Alzheimerțs DiseaseDocument32 pagesThe Dorsal Raphe Nucleus and Serotonin - Implications For Neuroplasticity Linked To Major Depression and Alzheimerțs DiseaseBogdan FedoNo ratings yet
- Neup 12143Document16 pagesNeup 12143Daniel Leonardo Guerrero BaqueroNo ratings yet
- Rūta Jurgaitienė Et Al - Morphometric Study of Age-Related Changes in The Human Intracardiac GangliaDocument8 pagesRūta Jurgaitienė Et Al - Morphometric Study of Age-Related Changes in The Human Intracardiac GangliaLuammmNo ratings yet
- Differences in Neural Connectivity Between The SubstantiaDocument5 pagesDifferences in Neural Connectivity Between The SubstantiaPamGarcíaNo ratings yet
- Sonic HedgehogDocument11 pagesSonic HedgehogPaola Rivera ÁvilaNo ratings yet
- FullDocument8 pagesFullrocambolescas perthNo ratings yet
- 1 Cambios RitmicosDocument16 pages1 Cambios RitmicosKevin RiofrioNo ratings yet
- Royal Society Proceedings: Biological SciencesDocument10 pagesRoyal Society Proceedings: Biological SciencesBipedalJoeNo ratings yet
- Craig and Banker 1994Document44 pagesCraig and Banker 1994Kanwal RashidNo ratings yet
- Histologia Del SNCDocument15 pagesHistologia Del SNCJuan Ignacio Cardenas RodriguezNo ratings yet
- Lavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexDocument8 pagesLavdas Et Al. - J.neurosc. - 1999 - The MGE Gives Rise To A Population of Early Neurons in The Developing Cerebral CortexCris AdFNo ratings yet
- Effect of Nerve Crush On Perikaryal Number and Volume of Neurons in Adult Rat Dorsal Root GanglionDocument7 pagesEffect of Nerve Crush On Perikaryal Number and Volume of Neurons in Adult Rat Dorsal Root Ganglionrnnr2159No ratings yet
- Hearing Research: Hisayuki Ojima, Kunio MurakamiDocument8 pagesHearing Research: Hisayuki Ojima, Kunio MurakamiFrontiersNo ratings yet
- New CellDocument60 pagesNew CellAna Paula CastroNo ratings yet
- Cookson 1996Document16 pagesCookson 1996M4shroomNo ratings yet
- 幹細胞與前瞻Document14 pages幹細胞與前瞻lunamedinaNo ratings yet
- Evidence For Neurogenesis in The LGDocument15 pagesEvidence For Neurogenesis in The LGUrielNo ratings yet
- Broadman 47 Dan AutisDocument4 pagesBroadman 47 Dan AutisSf AkhadiyatiNo ratings yet
- Mesopontine Organization of Cholinergic and Catecholaminergic Cell Groups in The Normal and Narcoleptic DogDocument13 pagesMesopontine Organization of Cholinergic and Catecholaminergic Cell Groups in The Normal and Narcoleptic DogAndjela RoganovicNo ratings yet
- DRG StimulationDocument11 pagesDRG StimulationShahnawaz ShahNo ratings yet
- Brain Functions Underlying SpeechDocument24 pagesBrain Functions Underlying SpeechHAKAN ÖZTEMÜRNo ratings yet
- Patterning The Developing DiencephalonDocument10 pagesPatterning The Developing DiencephalonAndres MoraNo ratings yet
- Ontology and NomenclatureDocument4 pagesOntology and NomenclatureCarlos Alb SeNo ratings yet
- Alcantara Et Al. - Cerebral Cortex - 2005 - BDNF-modulated Spatial Organization of CR Cells and GABAergic Neurons in The Marginal Zone Plays A Role in The Development of Cortical OrganizationDocument13 pagesAlcantara Et Al. - Cerebral Cortex - 2005 - BDNF-modulated Spatial Organization of CR Cells and GABAergic Neurons in The Marginal Zone Plays A Role in The Development of Cortical OrganizationCris AdFNo ratings yet
- Directing Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Document6 pagesDirecting Mouse Embryonic Neurosphere Differentiation Toward An Enriched Neuronal Population - Torrado 2014Rebeca ChávezNo ratings yet
- Fnana 08 00132Document10 pagesFnana 08 00132joaquimjsalvesNo ratings yet
- Genesis Avian Cells: Classic Embryonic Induction: The of Neural Crest ADocument6 pagesGenesis Avian Cells: Classic Embryonic Induction: The of Neural Crest ARANJAN KUMARNo ratings yet
- 1997 - Tsuji, Liberman - Intracellular Labeling of Auditory Nerve Fibers in Guinea Pig - Central and Peripheral ProjectionsDocument15 pages1997 - Tsuji, Liberman - Intracellular Labeling of Auditory Nerve Fibers in Guinea Pig - Central and Peripheral ProjectionsoverkindNo ratings yet
- Frenkel Exercise Protocol For Patient With Parkinsons DiseaseDocument39 pagesFrenkel Exercise Protocol For Patient With Parkinsons DiseaseHumaira Rahman0% (1)
- Cereb. Cortex 2003 Duan 950 61Document12 pagesCereb. Cortex 2003 Duan 950 61AlfonsoVillarroelCalistoNo ratings yet
- Study of Neurosecretory Cells in OdonataDocument16 pagesStudy of Neurosecretory Cells in Odonatashubhamatilkar04No ratings yet
- Olfactory Bulbectomy Induces Neuronal Rearrangement in The Entorhinal Cortex in The RatDocument7 pagesOlfactory Bulbectomy Induces Neuronal Rearrangement in The Entorhinal Cortex in The RatJesusNo ratings yet
- Hornung 2010Document14 pagesHornung 2010Leandy MartinezNo ratings yet
- Plexiform Neurofibroma of The Cauda Equina Presenting As Peroneal Muscular Atrophy (Muscle & Nerve, Vol. 19, Issue 2) (1996)Document4 pagesPlexiform Neurofibroma of The Cauda Equina Presenting As Peroneal Muscular Atrophy (Muscle & Nerve, Vol. 19, Issue 2) (1996)antonioopNo ratings yet
- Maturation of The Limbic System Revealed by MR FLAIR ImagingDocument5 pagesMaturation of The Limbic System Revealed by MR FLAIR ImagingTio Alf EugenioNo ratings yet
- 17-03 Fundamentos Neuroanatómicos Del LenguajeDocument27 pages17-03 Fundamentos Neuroanatómicos Del LenguajeAngela Solar San MartínNo ratings yet
- Neurogeneses Hipocampo 2013Document9 pagesNeurogeneses Hipocampo 2013Denise GarciaNo ratings yet
- Ebbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesDocument39 pagesEbbesson, Sven O. E. (1980) - Comparative Neurology of The Telencephalon. The Telencephalon of SnakesLivia PereiraNo ratings yet
- Jphysiol00385 0110Document9 pagesJphysiol00385 0110MichaelNo ratings yet
- Chapter 18 - Zebrafish Nervous Syste - 2020 - The Zebrafish in Biomedical ResearDocument9 pagesChapter 18 - Zebrafish Nervous Syste - 2020 - The Zebrafish in Biomedical ResearNicolas BaronNo ratings yet
- Cellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8From EverandCellular Dynamics of the Neuron: Symposia of the International Society for Cell Biology, Vol. 8No ratings yet
- Manuscript AtualizadoDocument22 pagesManuscript AtualizadomarioNo ratings yet
- Nucleus and Regions Density IntensityDocument4 pagesNucleus and Regions Density IntensitymarioNo ratings yet
- Manuscript AtualizadoDocument22 pagesManuscript AtualizadomarioNo ratings yet
- Manuscript AtualizadoDocument22 pagesManuscript AtualizadomarioNo ratings yet
- Nucleus and Regions Density IntensityDocument4 pagesNucleus and Regions Density IntensitymarioNo ratings yet
- Manuscript AtualizadoDocument22 pagesManuscript AtualizadomarioNo ratings yet
- Nucleus and Regions Density IntensityDocument4 pagesNucleus and Regions Density IntensitymarioNo ratings yet
- Nucleus and Regions Density IntensityDocument4 pagesNucleus and Regions Density IntensitymarioNo ratings yet
- BIOCHEMISTRY EXAMDocument7 pagesBIOCHEMISTRY EXAMchristinejoanNo ratings yet
- Biomarkers of Infection and SepsisDocument12 pagesBiomarkers of Infection and SepsisnuelitopNo ratings yet
- Finals 2nd Quiz Biochem MALANADocument3 pagesFinals 2nd Quiz Biochem MALANAJay-Ar MalanaNo ratings yet
- Cerebrospinal Fluid - AUBFDocument9 pagesCerebrospinal Fluid - AUBFMitch IbayNo ratings yet
- Neurotransmitter Dalam Fisiologi Saraf Otonom: Iwan Dwi Cahyono, Himawan Sasongko, Aria Dian PrimatikaDocument14 pagesNeurotransmitter Dalam Fisiologi Saraf Otonom: Iwan Dwi Cahyono, Himawan Sasongko, Aria Dian PrimatikaardianNo ratings yet
- 100 Drugs by Sayeed FinalDocument108 pages100 Drugs by Sayeed FinalAyeshik ChakrabortyNo ratings yet
- Regulation of Uterine Function: A Biochemical Conundrum in The Regulation of Smooth Muscle RelaxationDocument9 pagesRegulation of Uterine Function: A Biochemical Conundrum in The Regulation of Smooth Muscle Relaxationtiga sekawan sekawanNo ratings yet
- 3-Ascending Tracts of Spinal CordDocument28 pages3-Ascending Tracts of Spinal Cordmuhammad altaf100% (2)
- Primer: Frozen ShoulderDocument16 pagesPrimer: Frozen ShoulderJose PerezNo ratings yet
- Essentials of Psychiatric Mental Health Nursing 7th Edition Test BankDocument36 pagesEssentials of Psychiatric Mental Health Nursing 7th Edition Test Bankputvicaragewt788100% (39)
- Indiv Flow ChartDocument12 pagesIndiv Flow ChartDE LEON, CRONICA FAY G.No ratings yet
- Etm 21 3 9662 PDFDocument11 pagesEtm 21 3 9662 PDFachyNo ratings yet
- Research Paper 1 2Document12 pagesResearch Paper 1 2api-549249381No ratings yet
- Tissues and Cells Part 1 - Grade 7 - BiologyDocument4 pagesTissues and Cells Part 1 - Grade 7 - BiologyYasmina Jamal GholoumNo ratings yet
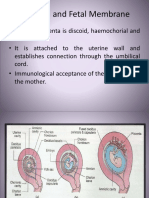
- Placenta and Fetal Membrane-6614Document45 pagesPlacenta and Fetal Membrane-6614Incredible DivineNo ratings yet
- How Body Systems Work TogetherDocument11 pagesHow Body Systems Work TogetherDonna RemitarNo ratings yet
- Presented By:-Samson S. Samuel 1 Year Guided By: - Dr. Fatima Saify Dr. Nidhi Sharma Dr. Kedarnath AgarwalDocument20 pagesPresented By:-Samson S. Samuel 1 Year Guided By: - Dr. Fatima Saify Dr. Nidhi Sharma Dr. Kedarnath AgarwalDrBhawna AroraNo ratings yet
- Prof. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineDocument50 pagesProf. N. Syabbalo: MB., CHB., PHD., FCCP., FRS., Fiba Professor of Physiology & MedicineHomeground entertainmentNo ratings yet
- Preparation of Artificial CSFDocument4 pagesPreparation of Artificial CSFThaarvena Retina0% (1)
- Primal Barbarian Paths ExplainedDocument9 pagesPrimal Barbarian Paths ExplainedMichael Taylor0% (1)
- Rodak Questions CompilationDocument40 pagesRodak Questions CompilationGertrudeshane IletoNo ratings yet
- Understanding Insect Life CyclesDocument38 pagesUnderstanding Insect Life CyclesYeye bonnellNo ratings yet
- Science: Modified Strategic Intervention MaterialsDocument24 pagesScience: Modified Strategic Intervention MaterialsRoxanne Manaloto100% (1)
- GR 10 Life Sciences IEB 3 in 1 Extracts PDFDocument16 pagesGR 10 Life Sciences IEB 3 in 1 Extracts PDFteeNo ratings yet
- Marma and Marma TherapyDocument14 pagesMarma and Marma TherapysunithaNo ratings yet
- Anatomy & Physiology (Chapter 1 - The Human Organism)Document23 pagesAnatomy & Physiology (Chapter 1 - The Human Organism)Ellie EileithyiaNo ratings yet
- The Interaction Between Autophagy, Helicobacter Pylori, and Gut Microbiota in Gastric CarcinogenesisDocument20 pagesThe Interaction Between Autophagy, Helicobacter Pylori, and Gut Microbiota in Gastric CarcinogenesisWaldo IzyNo ratings yet
- Developmental Biology GametogenesisDocument14 pagesDevelopmental Biology Gametogenesispragyamaharjan2No ratings yet
- Zen Masters TherapyDocument14 pagesZen Masters Therapysayyoni johriNo ratings yet
- Understanding the Cardiovascular SystemDocument7 pagesUnderstanding the Cardiovascular SystemKhrean Kae SantiagoNo ratings yet