You might also like
- 7.0 EnzymologyDocument7 pages7.0 EnzymologyHry WkNo ratings yet
- 1-Introduction To EnzymesDocument25 pages1-Introduction To EnzymesMarwa Mohamed AminNo ratings yet
- ENZYMESDocument16 pagesENZYMESWiza MulengaNo ratings yet
- Chapter 12 EnymesDocument6 pagesChapter 12 EnymesveymaramaNo ratings yet
- EnzymesDocument7 pagesEnzymesJuber mansuriNo ratings yet
- Enzymology: Dian Mulawarmanti School of Medicine Hang Tuah University 2006Document62 pagesEnzymology: Dian Mulawarmanti School of Medicine Hang Tuah University 2006dhika2496No ratings yet
- CH. 6 EnzymeDocument167 pagesCH. 6 EnzymeAyushi SinghalNo ratings yet
- EnzymesDocument20 pagesEnzymesDr.P.Natarajan100% (1)
- Enzymes in 40 CharactersDocument36 pagesEnzymes in 40 CharactersMilkiyas BirhanuNo ratings yet
- EnzymeDocument15 pagesEnzymeSheena Mhae LopeñaNo ratings yet
- EnzymesDocument37 pagesEnzymessxb4htnyhrNo ratings yet
- Enzymes: Catalysts of Life's ReactionsDocument20 pagesEnzymes: Catalysts of Life's ReactionsEniola JayeolaNo ratings yet
- ENZYME TECHNOLOGY: Biological Catalysts Speed Up Chemical ReactionsDocument131 pagesENZYME TECHNOLOGY: Biological Catalysts Speed Up Chemical ReactionsJanani RipplingrythmNo ratings yet
- Enzymes Mechanism of Enzyme ActionDocument6 pagesEnzymes Mechanism of Enzyme Actionkl42c4300No ratings yet
- EnzymesDocument28 pagesEnzymesGautamiNo ratings yet
- 1 Enzymes PDFDocument63 pages1 Enzymes PDFkunoz masrNo ratings yet
- EnzyDocument33 pagesEnzyPranaliNo ratings yet
- EnzymeDocument192 pagesEnzymeKishoreNo ratings yet
- Enzymes: Biological CatalystsDocument22 pagesEnzymes: Biological CatalystsRam Charan100% (1)
- Introduction to EnzymesDocument79 pagesIntroduction to EnzymesShafaqat Ghani Shafaqat GhaniNo ratings yet
- Enzymes ClassDocument152 pagesEnzymes ClassPrasthuthi Chowdary100% (2)
- Enzymes 12 5 19Document113 pagesEnzymes 12 5 19Arshad Mehmood MinhasNo ratings yet
- Enzymes and Proteins NotesDocument6 pagesEnzymes and Proteins NotesSarah MudaliarNo ratings yet
- Mr. Shardul S. WaghDocument24 pagesMr. Shardul S. WaghShardul WaghNo ratings yet
- Unit 7 EnzymesDocument85 pagesUnit 7 EnzymesAngelica Camille B. AbaoNo ratings yet
- EnzymesDocument5 pagesEnzymesAbhishek GangapurkarNo ratings yet
- Lecture 5 - Enzymes and CoenzymesDocument7 pagesLecture 5 - Enzymes and CoenzymesDoreenNo ratings yet
- Enzymes BN 2019Document67 pagesEnzymes BN 2019Rezan ThapaNo ratings yet
- Introduction to Medical BiochemistryDocument115 pagesIntroduction to Medical BiochemistryfikaduNo ratings yet
- Lectures 9 and 10, EnzymesDocument9 pagesLectures 9 and 10, Enzymesمجد محمودNo ratings yet
- Biology Grade 11 NotesDocument77 pagesBiology Grade 11 NotesHassenMohNo ratings yet
- Nomenclature and ClassificationDocument2 pagesNomenclature and ClassificationPoojaNo ratings yet
- Enzyme ClassificationDocument8 pagesEnzyme ClassificationR.KRISHNA KUMARINo ratings yet
- OUTLINE8Document8 pagesOUTLINE8Hjm ArNo ratings yet
- Copy of Copy of EnjymeDocument9 pagesCopy of Copy of Enjymeapi-327778003No ratings yet
- Enzymology To Kreb's CycleDocument34 pagesEnzymology To Kreb's Cycleferdinand padillaNo ratings yet
- UNIT 5 WorksheetDocument10 pagesUNIT 5 WorksheetlorNo ratings yet
- Enzymes AparajitaDocument49 pagesEnzymes Aparajitaappumohanty444No ratings yet
- BMS131 Lec05Document19 pagesBMS131 Lec05mariam tarekNo ratings yet
- Enzyme Structure and Mechanism of ActionDocument53 pagesEnzyme Structure and Mechanism of ActionAlok DhelditNo ratings yet
- Enzymes and Vitamins NotesDocument14 pagesEnzymes and Vitamins NotesRealyn ZambasNo ratings yet
- Module 3 ENZYMES Student's PDFDocument18 pagesModule 3 ENZYMES Student's PDFKerubin CastilloNo ratings yet
- CHY 47 EnzymesDocument143 pagesCHY 47 EnzymesElle BuhisanNo ratings yet
- Enzymes: Biological Catalysts That Speed Up ReactionsDocument34 pagesEnzymes: Biological Catalysts That Speed Up ReactionsShafaqat Ghani Shafaqat Ghani100% (2)
- Enzymes 1Document95 pagesEnzymes 1Hawi kelbessaNo ratings yet
- Enzymes and CofactorsDocument118 pagesEnzymes and Cofactorsur.yared21No ratings yet
- 4 - EnzymesDocument31 pages4 - EnzymescheckmateNo ratings yet
- EnzymesDocument7 pagesEnzymesKartik P. SahNo ratings yet
- What Is Enzyme?Document7 pagesWhat Is Enzyme?rentachin18No ratings yet
- Chapter 3 Enzymes Hormones VitaminsDocument133 pagesChapter 3 Enzymes Hormones VitaminsTran Danh NhanNo ratings yet
- Enzyme Technology by Martin F. ChaplinDocument149 pagesEnzyme Technology by Martin F. ChaplinAKSHAYNo ratings yet
- Unit - 3Document11 pagesUnit - 3Yitbarek TesfayeNo ratings yet
- Protein Biocatalyst (Enzyme)Document3 pagesProtein Biocatalyst (Enzyme)Khairul RidwanNo ratings yet
- Enzymes I: Definition, Structure, Properties, Nomenclature and ClassificationDocument15 pagesEnzymes I: Definition, Structure, Properties, Nomenclature and ClassificationHanan IsmailNo ratings yet
- Biological MoleculesDocument6 pagesBiological MoleculessanaullahNo ratings yet
- Biochemistry OralsDocument214 pagesBiochemistry Oralsdr.mumtaz09No ratings yet
- Enzyme and Vitamins, BiochemDocument2 pagesEnzyme and Vitamins, BiochemCraigyyNo ratings yet
- C5 EnzymesDocument19 pagesC5 EnzymesSoraya D. Al-ObinayNo ratings yet
- Unit 3 EnzymesDocument18 pagesUnit 3 Enzymeskamsjain6802No ratings yet
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- The Definition of Love - Andrew MarvellDocument1 pageThe Definition of Love - Andrew Marvellchrbacca100% (1)
- STDCSL Triangular-6Document3 pagesSTDCSL Triangular-6api-313618442No ratings yet
- How Red Blood Cells Transport Oxygen in the BodyDocument3 pagesHow Red Blood Cells Transport Oxygen in the BodySoha SonaNo ratings yet
- Blood-Borne Viruses in The Workplace: Guidance For Employers and EmployeesDocument7 pagesBlood-Borne Viruses in The Workplace: Guidance For Employers and EmployeesHUAWEI HUAWEINo ratings yet
- How Red Blood Cells Transport Oxygen in the BodyDocument3 pagesHow Red Blood Cells Transport Oxygen in the BodySoha SonaNo ratings yet
- Love of Life PDFDocument19 pagesLove of Life PDFTanyaLunaMoránNo ratings yet
- I Love PDF ConversionDocument6 pagesI Love PDF ConversionHUAWEI HUAWEINo ratings yet
- 2011 Blood TransfusionDocument6 pages2011 Blood TransfusionJeanne Mae Lee AfallaNo ratings yet
- Ucm 307643Document2 pagesUcm 307643HUAWEI HUAWEINo ratings yet
- Your Results For - Multiple Choice QuestionsDocument4 pagesYour Results For - Multiple Choice QuestionsHUAWEI HUAWEINo ratings yet
- 4 5979016688497068097Document3 pages4 5979016688497068097HUAWEI HUAWEINo ratings yet
- The Biochemical Changes During PregnancyDocument21 pagesThe Biochemical Changes During PregnancyHUAWEI HUAWEINo ratings yet
- The Scientific Research: Abdulla Salem Bin GhouthDocument20 pagesThe Scientific Research: Abdulla Salem Bin GhouthHUAWEI HUAWEINo ratings yet
- Minerals: Dr. Lotfi S. Bin Dahman Ph.D. M.D. Clinical Biochemistry HucomDocument7 pagesMinerals: Dr. Lotfi S. Bin Dahman Ph.D. M.D. Clinical Biochemistry HucomHUAWEI HUAWEINo ratings yet
- Scanned by CamscannerDocument3 pagesScanned by CamscannerHUAWEI HUAWEINo ratings yet
- Community-Based Education (CBE)Document10 pagesCommunity-Based Education (CBE)HUAWEI HUAWEINo ratings yet
- Biochem QbankDocument16 pagesBiochem Qbank786waqar786No ratings yet
- Review of NeurochemistryDocument18 pagesReview of NeurochemistryHUAWEI HUAWEINo ratings yet
- Common Medical Terms Sorted by ArabicDocument16 pagesCommon Medical Terms Sorted by ArabicAhmed El-Saadany100% (1)
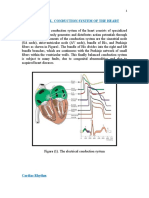
- Electrical Conduction System of the HeartDocument7 pagesElectrical Conduction System of the HeartHUAWEI HUAWEINo ratings yet
- الوحدة الخامسهDocument15 pagesالوحدة الخامسهHUAWEI HUAWEINo ratings yet
- أحياء ثانوية وزاري تجريبي2019 2020مDocument9 pagesأحياء ثانوية وزاري تجريبي2019 2020مHUAWEI HUAWEINo ratings yet
- States of Matter Test: NameDocument2 pagesStates of Matter Test: NameHUAWEI HUAWEINo ratings yet
- الوحدة الأولىDocument16 pagesالوحدة الأولىHUAWEI HUAWEINo ratings yet
- Test Bank 1Document8 pagesTest Bank 1IVLeeNo ratings yet
- Metabolism of Proteins: Dr. Lotfi S. Bin Dahman M.D. Ph.D. Clinical Biochemistry HucomDocument27 pagesMetabolism of Proteins: Dr. Lotfi S. Bin Dahman M.D. Ph.D. Clinical Biochemistry HucomHUAWEI HUAWEINo ratings yet
- كنترول أحياء فصل اولDocument6 pagesكنترول أحياء فصل اولHUAWEI HUAWEINo ratings yet
- Group Dynamics: Prof - Ali Batarfi MD, Ficms, Mhpe, MFPH (Uk)Document14 pagesGroup Dynamics: Prof - Ali Batarfi MD, Ficms, Mhpe, MFPH (Uk)HUAWEI HUAWEINo ratings yet
- الوحده الثالثهDocument8 pagesالوحده الثالثهHUAWEI HUAWEI100% (1)
- كنترول أحياء فصل اولDocument6 pagesكنترول أحياء فصل اولHUAWEI HUAWEINo ratings yet
- Thermosetting resins guide: Properties, applications and health risks of epoxy resinsDocument45 pagesThermosetting resins guide: Properties, applications and health risks of epoxy resinsDevendra100% (3)
- Poly (Vinyl Chloride) (PVC) Gaskets For Drain, Waste, and Vent (DWV), Sewer, Sanitary, and Storm Plumbing SystemsDocument3 pagesPoly (Vinyl Chloride) (PVC) Gaskets For Drain, Waste, and Vent (DWV), Sewer, Sanitary, and Storm Plumbing SystemsveintavoNo ratings yet
- Carbonyl Compund Subjective QuestionsDocument11 pagesCarbonyl Compund Subjective QuestionsVinod AgrawalNo ratings yet
- Review Article Bougainvillea Genus: A Review On PhytochemistryDocument18 pagesReview Article Bougainvillea Genus: A Review On PhytochemistryANA DANIELA FIGUEROA HERNANDEZNo ratings yet
- The Role of ATP in Metabolism: Mutiara Dewi ListiyanawatiDocument21 pagesThe Role of ATP in Metabolism: Mutiara Dewi ListiyanawatiTurasno Abi ZahraNo ratings yet
- June 10, 1958: Continuous Hydrolysis of EpichlorohydrinDocument4 pagesJune 10, 1958: Continuous Hydrolysis of EpichlorohydrinAbdur Rochim Al IchwanNo ratings yet
- MALACHITE GREEN SAFETY DATA SHEETDocument9 pagesMALACHITE GREEN SAFETY DATA SHEETRomero GhyoNo ratings yet
- Class XII Chemistry 23-24 Pre-Board SolDocument14 pagesClass XII Chemistry 23-24 Pre-Board Solap1124214No ratings yet
- DINDA - Biochem Pract ReportDocument18 pagesDINDA - Biochem Pract ReportdindaNo ratings yet
- Article WJPR 1513745964Document12 pagesArticle WJPR 1513745964Sivakumar LakshminarayananNo ratings yet
- 099-106 Sample Cells MC PDFDocument8 pages099-106 Sample Cells MC PDFluis miguel huarita castellonNo ratings yet
- Process Synthesis and Optimization of Propylene - Propane Separation Using Vapor Recompression and Self-Heat RecuperationDocument8 pagesProcess Synthesis and Optimization of Propylene - Propane Separation Using Vapor Recompression and Self-Heat RecuperationForcus onNo ratings yet
- Analysis of Fatty Acid Methyl Esters in Edible Oils: Application Note - An172Document8 pagesAnalysis of Fatty Acid Methyl Esters in Edible Oils: Application Note - An172salvaleuvenNo ratings yet
- Cambridge IGCSE™: Agriculture 0600/11 October/November 2020Document25 pagesCambridge IGCSE™: Agriculture 0600/11 October/November 2020Sraboni ChowdhuryNo ratings yet
- Satellite-Detected Early Summer Coccolithophore Blooms and Their Interannual Variability in The Black SeaDocument15 pagesSatellite-Detected Early Summer Coccolithophore Blooms and Their Interannual Variability in The Black SeaTÜLAY ÇOKACARNo ratings yet
- Información Nutricional Citrato de Magnesio Food PSTDocument2 pagesInformación Nutricional Citrato de Magnesio Food PSTPamela Urrea AtenasNo ratings yet
- SEO-Optimized Catalog of Medical UniformsDocument36 pagesSEO-Optimized Catalog of Medical UniformsReza Yuni SandraNo ratings yet
- Sodium Bicarbonate As Inorganic Carbon Source For Higher - 2016 - AchievementsDocument7 pagesSodium Bicarbonate As Inorganic Carbon Source For Higher - 2016 - Achievementssameer sahaanNo ratings yet
- Efficacy of Papaya Seeds (OUTLINE) 2.1Document12 pagesEfficacy of Papaya Seeds (OUTLINE) 2.1wynferd ssNo ratings yet
- Org Chem Final ReviewerDocument7 pagesOrg Chem Final ReviewerblessaNo ratings yet
- 10 - The Genetic Code and Translation Revised 9-24-2018Document16 pages10 - The Genetic Code and Translation Revised 9-24-2018Sheraz ChaudharyNo ratings yet
- St. Andrew's Junior College H2 Chemistry 2013 Organic Chemistry Lecture Notes 5 ArenesDocument26 pagesSt. Andrew's Junior College H2 Chemistry 2013 Organic Chemistry Lecture Notes 5 ArenesArvin LiangdyNo ratings yet
- Palladium-Catalyzed Conversion of Aryl and Vinyl Triflates To Bromides and ChloridesDocument3 pagesPalladium-Catalyzed Conversion of Aryl and Vinyl Triflates To Bromides and ChloridesnobodycNo ratings yet
- Progress and Challenges For Replacing N Methyl 2 Py 2024 Progress in NaturalDocument13 pagesProgress and Challenges For Replacing N Methyl 2 Py 2024 Progress in Naturalmosab.backkupNo ratings yet
- Mechanical Properties of Composites Using Natural Rubber With Epoxy ResinDocument106 pagesMechanical Properties of Composites Using Natural Rubber With Epoxy ResinLeksir KamiliaNo ratings yet
- Thioplast G4: Liquid Polysulfide Polymer With Thiol End Groups (MW 1800)Document3 pagesThioplast G4: Liquid Polysulfide Polymer With Thiol End Groups (MW 1800)kebenshanNo ratings yet
- JPAM Vol 15 Issue1 P 285-299Document15 pagesJPAM Vol 15 Issue1 P 285-299Jefri Nur HidayatNo ratings yet
- KA-Modern Analyitical Chreymistry - David Harvey - 1Document11 pagesKA-Modern Analyitical Chreymistry - David Harvey - 1Kallolona SidrapNo ratings yet
- Role of biotechnology in food processing and preservationDocument51 pagesRole of biotechnology in food processing and preservationrani padamavathiNo ratings yet
- Chemistry - Are Allenes Optically Active - QuoraDocument3 pagesChemistry - Are Allenes Optically Active - QuoraAhmadulhaqNo ratings yet