You might also like
- Mesothelioma: From Research to Clinical PracticeFrom EverandMesothelioma: From Research to Clinical PracticeGiovanni Luca CeresoliNo ratings yet
- Atlas of Mohs and Frozen Section Cutaneous PathologyFrom EverandAtlas of Mohs and Frozen Section Cutaneous PathologyMichael B. MorganNo ratings yet
- VitiligoDocument6 pagesVitiligohukaNo ratings yet
- Vitiligo: Symptoms, Pathogenesis and TreatmentDocument6 pagesVitiligo: Symptoms, Pathogenesis and TreatmentdincajeanNo ratings yet
- Investigationoftheefficacyandtolerabilityof DR Michaelsalsobrandedas Eczitinexand Itchinex EczitinextopicalproductsinthetreatmentofatopicdermatitisinchildrenDocument10 pagesInvestigationoftheefficacyandtolerabilityof DR Michaelsalsobrandedas Eczitinexand Itchinex EczitinextopicalproductsinthetreatmentofatopicdermatitisinchildrenLuisNo ratings yet
- Extracellular Matrix Remodeling in Equine Sarcoid PDFDocument11 pagesExtracellular Matrix Remodeling in Equine Sarcoid PDFSatriaNo ratings yet
- Med Res Rev 20173798-148Document52 pagesMed Res Rev 20173798-148Estefania VelascoNo ratings yet
- Fluorescence Biomodulation in The Management of AcDocument7 pagesFluorescence Biomodulation in The Management of AcAnisaNo ratings yet
- Thymol Jounal 2Document22 pagesThymol Jounal 2asmidakamarudinNo ratings yet
- EuropeanpaperDocument11 pagesEuropeanpaperMostafa EidNo ratings yet
- AEuropeanprospectiverandomizedplacebo controlleddoubleblindStudyontheefficacyandsafetyofDrMichaelsalsobrandedasSoratinexproductfamilyforstablechronicplaquepsoriasisDocument7 pagesAEuropeanprospectiverandomizedplacebo controlleddoubleblindStudyontheefficacyandsafetyofDrMichaelsalsobrandedasSoratinexproductfamilyforstablechronicplaquepsoriasisLuisNo ratings yet
- What Is The Importance of Demodex Folliculorum in Behçet's Disease?Document6 pagesWhat Is The Importance of Demodex Folliculorum in Behçet's Disease?Ghea Tahara SugiantoNo ratings yet
- Nomura 2020Document13 pagesNomura 2020npp52kxzfyNo ratings yet
- Clinics OncologyDocument5 pagesClinics OncologyClinics OncologyNo ratings yet
- Sundaram Et Al-2018-The FEBS JournalDocument20 pagesSundaram Et Al-2018-The FEBS JournalYusroqNo ratings yet
- Fcell 07 00143Document13 pagesFcell 07 00143Tara KamaranNo ratings yet
- Anil Kumar (Cell Biology) - 1Document11 pagesAnil Kumar (Cell Biology) - 1BISWANATH ROUTNo ratings yet
- Artigo argilaBOMMMDocument22 pagesArtigo argilaBOMMMAdriana SampaioNo ratings yet
- Deka Upl Melasma Ipl Zoccali Et Al 2010Document9 pagesDeka Upl Melasma Ipl Zoccali Et Al 2010isma drNo ratings yet
- Necrotizing Fasciitis Caused by Haemophilus InflueDocument5 pagesNecrotizing Fasciitis Caused by Haemophilus InflueAgussalim SaniNo ratings yet
- Fighting Against Skin Aging: The Way From Bench To Bedside: Shoubing Zhang and Enkui DuanDocument10 pagesFighting Against Skin Aging: The Way From Bench To Bedside: Shoubing Zhang and Enkui DuanRiley RilanNo ratings yet
- Fimosis JournalDocument4 pagesFimosis JournalMegawaty Putri SyaefulNo ratings yet
- The Role of Autophagy in Skin Fibroblasts, Keratinocytes, Melanocytes, and Epidermal Stem CellsDocument7 pagesThe Role of Autophagy in Skin Fibroblasts, Keratinocytes, Melanocytes, and Epidermal Stem Cellsvanessa_werbickyNo ratings yet
- Photoderm Photoimm Photomed - 2021 - Krutmann - Daily Photoprotection To Prevent PhotoagingDocument8 pagesPhotoderm Photoimm Photomed - 2021 - Krutmann - Daily Photoprotection To Prevent PhotoagingDra. Marcela Catalina Fandiño VargasNo ratings yet
- Phenotypic, Functional, and Metabolic Heterogeneity of Immune Cells Infiltrating Non Small Cell Lung CancerDocument14 pagesPhenotypic, Functional, and Metabolic Heterogeneity of Immune Cells Infiltrating Non Small Cell Lung CancerWilliamRayCassidyNo ratings yet
- Physiopathology of Melasma PDFDocument16 pagesPhysiopathology of Melasma PDFKenneth MartinoNo ratings yet
- Antimalarial Potential, LC-MS Secondary Metabolite Profiling and Computational Studies of Zingiber OfficinaleDocument18 pagesAntimalarial Potential, LC-MS Secondary Metabolite Profiling and Computational Studies of Zingiber Officinale2281246No ratings yet
- Microorganisms 08 01752Document32 pagesMicroorganisms 08 01752Mariel IslasNo ratings yet
- Clinico Morphological Evaluation of Melasma and Other Hyperpigmentary Disorders of SkinDocument25 pagesClinico Morphological Evaluation of Melasma and Other Hyperpigmentary Disorders of SkinBaru Chandrasekhar RaoNo ratings yet
- Pigment Int EngimaofcolorintineaversicolorDocument5 pagesPigment Int EngimaofcolorintineaversicolorKuldeepSinghBanaNo ratings yet
- Autophagy, Anoikis, Ferroptosis, Necroptosis, and Endoplasmic Reticulum Stress - Potential Applications in Melanoma TherapyDocument9 pagesAutophagy, Anoikis, Ferroptosis, Necroptosis, and Endoplasmic Reticulum Stress - Potential Applications in Melanoma TherapyEstrella BenitoNo ratings yet
- PCMR 12404Document14 pagesPCMR 12404Riefka Ananda ZulfaNo ratings yet
- Ijms 22 03035 v2Document3 pagesIjms 22 03035 v2Winona YuvaNo ratings yet
- Granuloma PCLDocument5 pagesGranuloma PCLEmrys1987No ratings yet
- TaiwanJOphthalmol000-3647805 100758Document11 pagesTaiwanJOphthalmol000-3647805 100758arnanda raizhaNo ratings yet
- Front Immunol PrintedDocument17 pagesFront Immunol PrintedMELANI BELLO FLORESNo ratings yet
- Ghafourian Et Al 2014 Vitiligo Symptoms Pathogenesis and TreatmentDocument5 pagesGhafourian Et Al 2014 Vitiligo Symptoms Pathogenesis and Treatmentnramrattan475No ratings yet
- Polymers 2021Document17 pagesPolymers 2021Oana MariaNo ratings yet
- Ma 2024 Biomed. Mater. 19 015008Document13 pagesMa 2024 Biomed. Mater. 19 015008asyaroesNo ratings yet
- FlyerDocument3 pagesFlyerTerem TebautaNo ratings yet
- Mechanism of Induction of Fibroblast To Corneal Endothelial CellDocument4 pagesMechanism of Induction of Fibroblast To Corneal Endothelial CelltinaNo ratings yet
- Ah SkinDocument24 pagesAh SkinJuliana BrunettoNo ratings yet
- Biomedicines 09 00435 v2Document19 pagesBiomedicines 09 00435 v2Jesus Vargas GalindoNo ratings yet
- Fgene 11 579738Document11 pagesFgene 11 579738Gustavo OrtízNo ratings yet
- Admin,+27 Ism.v11i1.633Document5 pagesAdmin,+27 Ism.v11i1.633Fasih Djam'an DikaNo ratings yet
- Journal of Pineal Research - 2017 - Mayo - Melatonin and Sirtuins A Not So Unexpected RelationshipDocument17 pagesJournal of Pineal Research - 2017 - Mayo - Melatonin and Sirtuins A Not So Unexpected RelationshipBrenda AkemiNo ratings yet
- Cells 12 02483Document20 pagesCells 12 02483keerthi.sakthi0794No ratings yet
- Oculocutaneous Albinism and Cutaneous MaDocument9 pagesOculocutaneous Albinism and Cutaneous MaFrancialNo ratings yet
- Mesenchymal Stem Cell-Conditioned Medium (Secretome) in Skin Aging A Systematic ReviewDocument24 pagesMesenchymal Stem Cell-Conditioned Medium (Secretome) in Skin Aging A Systematic ReviewThomas UtomoNo ratings yet
- Martignago Et Al-2019-Lasers in Medical ScienceDocument9 pagesMartignago Et Al-2019-Lasers in Medical Sciencedevrajsinhchauhan8055No ratings yet
- New CarcinogenicDocument18 pagesNew CarcinogenicMelati HasnailNo ratings yet
- Cancers 14 00476Document25 pagesCancers 14 00476Christine OlivarNo ratings yet
- Photodyn Ther 2013 Novak Et AlDocument17 pagesPhotodyn Ther 2013 Novak Et Alenglish-exactlyNo ratings yet
- Histopathological Variations in Ameloblastoma - Cases in A Tertiary Care Center of NepalDocument7 pagesHistopathological Variations in Ameloblastoma - Cases in A Tertiary Care Center of NepalDinesh YadavNo ratings yet
- The Influence of Cellular Seeding Density in The MDocument11 pagesThe Influence of Cellular Seeding Density in The Mcarolina mejiaNo ratings yet
- Structure Modification of Ethyl P-Methoxycinnamate and Their Bioassay As Chemopreventive Agent Against Mice's FibrosarcomaDocument6 pagesStructure Modification of Ethyl P-Methoxycinnamate and Their Bioassay As Chemopreventive Agent Against Mice's FibrosarcomaFATTANo ratings yet
- AntioxidantesDocument7 pagesAntioxidantesSilvana OliveiraNo ratings yet
- TH 8Document15 pagesTH 8SnoAbuAlSamedNo ratings yet
- Hamzah Noegraha - Jurnal Tugas 3 Biofilm MikrobologiDocument7 pagesHamzah Noegraha - Jurnal Tugas 3 Biofilm MikrobologiHamzah NoegrahaNo ratings yet
- Role of Peeling in HPIDocument8 pagesRole of Peeling in HPIirwan junNo ratings yet
- Jurnal Efficacy and Tolerability of Adjunctive Lacosamide in Pediatric Patients With Focal SeizuresDocument15 pagesJurnal Efficacy and Tolerability of Adjunctive Lacosamide in Pediatric Patients With Focal SeizuresAnida HasnaNo ratings yet
- 0256-4947 2017 449Document6 pages0256-4947 2017 449vanessa armeliaNo ratings yet
- Tuberculous MeningoencephalitisDocument3 pagesTuberculous MeningoencephalitisAnida HasnaNo ratings yet
- TRD 82 E20Document9 pagesTRD 82 E20Anida HasnaNo ratings yet
- Therapy Studies What Question Did The Study Ask?Document2 pagesTherapy Studies What Question Did The Study Ask?Apt RiskaNo ratings yet
- Jurnal Causes and Factors Associated With Neonatal Seizure and Its Short-Term Outcome A Retrospective Prognostic Cohort StudyDocument10 pagesJurnal Causes and Factors Associated With Neonatal Seizure and Its Short-Term Outcome A Retrospective Prognostic Cohort StudyAnida HasnaNo ratings yet
- Efficacy of Artemether Lumefantrine Versus DihydroartemisininDocument8 pagesEfficacy of Artemether Lumefantrine Versus DihydroartemisininAnida HasnaNo ratings yet
- Jurnal Randomized Phase 2 Study of Adjunctive Cenobamate in Patients With Uncontrolled Focal SeizuresDocument12 pagesJurnal Randomized Phase 2 Study of Adjunctive Cenobamate in Patients With Uncontrolled Focal SeizuresAnida HasnaNo ratings yet
- Review Article: Tuberculous Meningitis: Diagnosis and Treatment OverviewDocument10 pagesReview Article: Tuberculous Meningitis: Diagnosis and Treatment OverviewMeldaNo ratings yet
- BMJ f360 FullDocument15 pagesBMJ f360 FullAnida HasnaNo ratings yet
- 3994 11816 1 SMDocument8 pages3994 11816 1 SMRahmi Fitri SyamNo ratings yet
- 3994 11816 1 SMDocument8 pages3994 11816 1 SMRahmi Fitri SyamNo ratings yet
- Tuberculous Meningoencephalitis: 2. Case Pre-SentationDocument2 pagesTuberculous Meningoencephalitis: 2. Case Pre-SentationAnida HasnaNo ratings yet
- CERa1 FSL PF1AG SHU PNSL1Document3 pagesCERa1 FSL PF1AG SHU PNSL1Anida HasnaNo ratings yet
- Actaapa 2020 19Document3 pagesActaapa 2020 19iqbal dsNo ratings yet
- DMCN 14015Document10 pagesDMCN 14015Anida HasnaNo ratings yet
- The Role of Sunscreen in Melasma and Postinflammatory Hyperpigmentation - (PAUTHORS), Indian Journal of DermatologyDocument6 pagesThe Role of Sunscreen in Melasma and Postinflammatory Hyperpigmentation - (PAUTHORS), Indian Journal of DermatologyAnida HasnaNo ratings yet
- Journal of The American Academy of Dermatology Volume 77 Issue 4 2017 (Doi 10.1016 - J.jaad.2017.01.036) Chaowattanapanit, Suteeraporn Silpa-Archa, Narumol Kohli, Inde - Postinflammatory HyperpigmeDocument15 pagesJournal of The American Academy of Dermatology Volume 77 Issue 4 2017 (Doi 10.1016 - J.jaad.2017.01.036) Chaowattanapanit, Suteeraporn Silpa-Archa, Narumol Kohli, Inde - Postinflammatory HyperpigmedikativiNo ratings yet
- Clinical Analysis, Diagnosis, and Treatment of A 48-Year-Old Female Filipino With Riehl MelanosisDocument9 pagesClinical Analysis, Diagnosis, and Treatment of A 48-Year-Old Female Filipino With Riehl Melanosisiqbal dsNo ratings yet
- 3994 11816 1 SMDocument8 pages3994 11816 1 SMRahmi Fitri SyamNo ratings yet
- 3994 11816 1 SMDocument8 pages3994 11816 1 SMRahmi Fitri SyamNo ratings yet
- Histopstological Features of Riehl MelanosisDocument5 pagesHistopstological Features of Riehl MelanosisAnida HasnaNo ratings yet
- 1 Silpaarcha 2017Document15 pages1 Silpaarcha 2017lucianaNo ratings yet
- PIH in Dark Skin DR Abirami Pararajasingam and DR Sandeep CliffeDocument4 pagesPIH in Dark Skin DR Abirami Pararajasingam and DR Sandeep CliffeAnida HasnaNo ratings yet
- 2014 Article 639Document8 pages2014 Article 639Anida HasnaNo ratings yet
- Jurnal Gizi Buruk BalitaDocument9 pagesJurnal Gizi Buruk BalitaDokter Daphine SatriaNo ratings yet
- Applsci 10 02049Document10 pagesApplsci 10 02049Anida HasnaNo ratings yet
- PIH Model Mousein PCMR 2020Document12 pagesPIH Model Mousein PCMR 2020Anida HasnaNo ratings yet
- Fitzgerald 2020 American College of Rheumatology GDocument18 pagesFitzgerald 2020 American College of Rheumatology GAnida HasnaNo ratings yet
- 2020 Article 479Document5 pages2020 Article 479Anida HasnaNo ratings yet
- Tatanka Oyate 2 EngDocument3 pagesTatanka Oyate 2 EngdesiNo ratings yet
- Manual Murray OkDocument52 pagesManual Murray OkGIOVANNINo ratings yet
- Torres v. People (2011)Document2 pagesTorres v. People (2011)Gigi LucilleNo ratings yet
- Forrester Hybrid Integration PlatformDocument17 pagesForrester Hybrid Integration PlatformrtNo ratings yet
- Colin Campbell - Interview The Lady in WhiteDocument1 pageColin Campbell - Interview The Lady in WhiteMarcelo OlmedoNo ratings yet
- Adult Student Boundaries BrochureDocument2 pagesAdult Student Boundaries BrochuretalatalamarkyaldrinNo ratings yet
- Bupa Statement To ABCDocument1 pageBupa Statement To ABCABC News OnlineNo ratings yet
- Intermediate Macroeconomics Sec 222Document163 pagesIntermediate Macroeconomics Sec 222Katunga MwiyaNo ratings yet
- English IODocument6 pagesEnglish IOAarav MathurNo ratings yet
- IPCC - Fast Track Accounting - 35e PDFDocument37 pagesIPCC - Fast Track Accounting - 35e PDFRam IyerNo ratings yet
- Module 6:market Segmentation, Market Targeting, and Product PositioningDocument16 pagesModule 6:market Segmentation, Market Targeting, and Product Positioningjanel anne yvette sorianoNo ratings yet
- Antenatal Assessment of Fetal Well Being FileminimizerDocument40 pagesAntenatal Assessment of Fetal Well Being FileminimizerPranshu Prajyot 67No ratings yet
- The List of the Korean Companies in the UAE: Company Name 이름 Off. Tel Off. Fax P.O.BoxDocument34 pagesThe List of the Korean Companies in the UAE: Company Name 이름 Off. Tel Off. Fax P.O.Boxnguyen phuong anh100% (1)
- Walt Disney OutlineDocument9 pagesWalt Disney Outlineapi-234693246No ratings yet
- Analisa Getaran BerlebihDocument5 pagesAnalisa Getaran BerlebihMatsaid ReksonoNo ratings yet
- Carbon Trading-The Future Money Venture For IndiaDocument11 pagesCarbon Trading-The Future Money Venture For IndiaijsretNo ratings yet
- Line Follower Robot Using ArduinoDocument5 pagesLine Follower Robot Using Arduinochockalingam athilingam100% (1)
- Pamrex - Manhole CoversDocument6 pagesPamrex - Manhole CoversZivadin LukicNo ratings yet
- Soalan Tugasan HBMT2103 - V2 Sem Mei 2015Document10 pagesSoalan Tugasan HBMT2103 - V2 Sem Mei 2015Anonymous wgrNJjANo ratings yet
- Jewish Standard, December 18, 2015Document64 pagesJewish Standard, December 18, 2015New Jersey Jewish StandardNo ratings yet
- Magness - The Tomb of Jesus and His Family - Exploring Ancient Jewish Tombs Near Jerusalem's Walls Book ReviewDocument5 pagesMagness - The Tomb of Jesus and His Family - Exploring Ancient Jewish Tombs Near Jerusalem's Walls Book Reviewarbg100% (1)
- Case Study of Hyper Loop TrainDocument3 pagesCase Study of Hyper Loop TrainkshitijNo ratings yet
- Operating BudgetDocument38 pagesOperating BudgetRidwan O'connerNo ratings yet
- BSBPEF501: Manage Personal and Professional DevelopmentDocument88 pagesBSBPEF501: Manage Personal and Professional DevelopmentBruce100% (1)
- Employer BrandingDocument17 pagesEmployer BrandingSushri PadhiNo ratings yet
- 6426c-Enu Trainerhandbook 2Document392 pages6426c-Enu Trainerhandbook 2mu771_No ratings yet
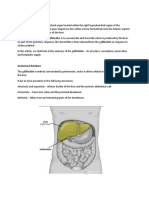
- AnatomyDocument4 pagesAnatomySureen PaduaNo ratings yet
- Civil TutorialDocument638 pagesCivil TutorialEmmanuel Mends FynnNo ratings yet
- Polaris Ranger 500 ManualDocument105 pagesPolaris Ranger 500 ManualDennis aNo ratings yet
- ASVAB Core ReviewDocument208 pagesASVAB Core Reviewstanley100% (1)