You might also like
- The Myth of Autism: How a Misunderstood Epidemic Is Destroying Our ChildrenFrom EverandThe Myth of Autism: How a Misunderstood Epidemic Is Destroying Our ChildrenRating: 2.5 out of 5 stars2.5/5 (8)
- Full Download Biology of Humans Concepts Applications and Issues 6th Edition Goodenough Solutions ManualDocument36 pagesFull Download Biology of Humans Concepts Applications and Issues 6th Edition Goodenough Solutions Manualbrumfieldridleyvip97% (35)
- Lewy Body Dementia SlidesDocument111 pagesLewy Body Dementia SlidesDilini de Silva-ChenNo ratings yet
- MycoViro - Complete Handouts (AMCC)Document46 pagesMycoViro - Complete Handouts (AMCC)Martin Clyde100% (1)
- Approach To A Patient With AtaxiaDocument66 pagesApproach To A Patient With AtaxiaBobeică S MihailNo ratings yet
- Bio1 11 - 12 Q1 0103 PF FDDocument35 pagesBio1 11 - 12 Q1 0103 PF FDRobbie Jule De Los SantosNo ratings yet
- Nutrition in Islamic PerspectiveDocument15 pagesNutrition in Islamic PerspectiveRangga Alam VaneoNo ratings yet
- Histopath Easy ReviewerDocument35 pagesHistopath Easy ReviewerSamantha Samarita100% (4)
- 2009 A Level H2 Biology P2 AnsDocument11 pages2009 A Level H2 Biology P2 AnsjoannetzyNo ratings yet
- Uner Tan Syndrome: History, Clinical Evaluations, Genetics, and The Dynamics of Human QuadrupedalismDocument12 pagesUner Tan Syndrome: History, Clinical Evaluations, Genetics, and The Dynamics of Human QuadrupedalismFrontiersNo ratings yet
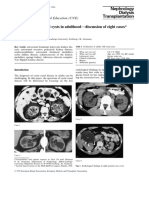
- VACTERL-Hydrocephaly, DK-Phocomelia, and Cerebro-Cardio-Radio-Reno-Rectal CommunityDocument6 pagesVACTERL-Hydrocephaly, DK-Phocomelia, and Cerebro-Cardio-Radio-Reno-Rectal CommunitySarly FebrianaNo ratings yet
- Fearnley Lees BRAIN 1991Document19 pagesFearnley Lees BRAIN 1991Sam StuartNo ratings yet
- Pallister-Killian Mosaic Syndrome in An Omani NewbDocument5 pagesPallister-Killian Mosaic Syndrome in An Omani NewbFrank Harry LampardNo ratings yet
- Krabbe's ImagingDocument4 pagesKrabbe's Imagingshruti kumarNo ratings yet
- Familial Neuronal Migration Disorder: Subcortical: Laminar Heterotopia in A Mother and Pachygyria in The SonDocument4 pagesFamilial Neuronal Migration Disorder: Subcortical: Laminar Heterotopia in A Mother and Pachygyria in The SoncthjgfytvfnoNo ratings yet
- A New Familial Adult-Onset Leukodystrophy Manifesting As Cerebellar Ataxia and DementiaDocument9 pagesA New Familial Adult-Onset Leukodystrophy Manifesting As Cerebellar Ataxia and DementiaKristineNo ratings yet
- Loss of Function of KIAA2022 Causes Mild To Severe Intellectual Disability With An Autism Spectrum Disorder and Impairs Neurite OutgrowthDocument9 pagesLoss of Function of KIAA2022 Causes Mild To Severe Intellectual Disability With An Autism Spectrum Disorder and Impairs Neurite OutgrowthFrontiersNo ratings yet
- Brain 2014 Yu Brain Awu239Document5 pagesBrain 2014 Yu Brain Awu239Angeles RbNo ratings yet
- Nob 30093Document7 pagesNob 30093Bilel Le ChevalierNo ratings yet
- IJHG 09 2 105 09 355 Vashist M TTDocument3 pagesIJHG 09 2 105 09 355 Vashist M TTcindyNo ratings yet
- Confirmation of The Catania Brachydactylous Type of Acrofacial DysostosisDocument4 pagesConfirmation of The Catania Brachydactylous Type of Acrofacial DysostosisSergioFernandezNo ratings yet
- Microduplication of xp22.31Document4 pagesMicroduplication of xp22.31Roberth SanchezNo ratings yet
- Brain Imaging in A Case of Capgras' SyndromeDocument6 pagesBrain Imaging in A Case of Capgras' SyndromeHusain RezaNo ratings yet
- Agenesia CerebeloDocument5 pagesAgenesia CerebeloDiretoria CientíficaNo ratings yet
- Wieczorek 2003Document5 pagesWieczorek 2003ladybieibiNo ratings yet
- Prenatal Diagnosis - 2004 - Pajkrt - Fetal Cardiac Anomalies and Genetic SyndromesDocument12 pagesPrenatal Diagnosis - 2004 - Pajkrt - Fetal Cardiac Anomalies and Genetic SyndromesGabi ONo ratings yet
- Letter To The Editor: Autism and GeneticsDocument2 pagesLetter To The Editor: Autism and GeneticsHarshit AmbeshNo ratings yet
- Broadman 47 Dan AutisDocument4 pagesBroadman 47 Dan AutisSf AkhadiyatiNo ratings yet
- Shapiro 1997Document6 pagesShapiro 1997JohnnyNo ratings yet
- Schnecken Becken DysplasiaDocument4 pagesSchnecken Becken Dysplasiadr sutjiNo ratings yet
- Genetics of Human Heterotaxias: ReviewDocument9 pagesGenetics of Human Heterotaxias: ReviewJacob Trisusilo SaleanNo ratings yet
- Leukodystrophies and MucopolysacridosisDocument78 pagesLeukodystrophies and MucopolysacridosisMobin Ur Rehman KhanNo ratings yet
- Yamamoto 1984Document2 pagesYamamoto 1984Victor Lopez SueroNo ratings yet
- The Basal Ganglia - Clinical CorrelatesDocument4 pagesThe Basal Ganglia - Clinical Correlatesalecsandra96No ratings yet
- HHS Public Access: XRCC1 Mutation Is Associated With PARP1 Hyperactivation andDocument25 pagesHHS Public Access: XRCC1 Mutation Is Associated With PARP1 Hyperactivation andThaís MesequeNo ratings yet
- Band Heterotopia in Zellweger Syndrome (Cerebro-Hepato Renal Syndrome)Document1 pageBand Heterotopia in Zellweger Syndrome (Cerebro-Hepato Renal Syndrome)ZazzZaffaNo ratings yet
- The Cerebellum - Clinical CorrelatesDocument5 pagesThe Cerebellum - Clinical Correlatesalecsandra96No ratings yet
- Ejhg 2013130Document5 pagesEjhg 2013130Flavia ChNo ratings yet
- Prevalence of Capitellar Osteochondritis Dissecans in Children With Chronic Radial Head Subluxation and DislocationDocument7 pagesPrevalence of Capitellar Osteochondritis Dissecans in Children With Chronic Radial Head Subluxation and DislocationAb Hakim MuslimNo ratings yet
- Effect Ofl Dopa On Oculogyric Crises inDocument2 pagesEffect Ofl Dopa On Oculogyric Crises inJoséNo ratings yet
- Carrozzo 1997Document6 pagesCarrozzo 1997special spamailNo ratings yet
- Progressive Increase of The Mutated Mitochondria1 DNA Fraction in Kearns-Sayre SyndromeDocument6 pagesProgressive Increase of The Mutated Mitochondria1 DNA Fraction in Kearns-Sayre SyndromeGréta BotyánszkiNo ratings yet
- PAPER (ENG) - (Damasio A. R., 1983) The Anatomic Basis of Pure AlexiaDocument12 pagesPAPER (ENG) - (Damasio A. R., 1983) The Anatomic Basis of Pure AlexiaAldo Hip NaranjoNo ratings yet
- Posterior Cortical Dementia: Lost But Not ForgettingDocument2 pagesPosterior Cortical Dementia: Lost But Not ForgettingElisabet GobelliNo ratings yet
- RJN 2021 4 Art-18Document4 pagesRJN 2021 4 Art-18Emilia LunguNo ratings yet
- Plugin Ibnt04i4p383Document2 pagesPlugin Ibnt04i4p383bobiuuuuuNo ratings yet
- Subacute Sclerosing Panencephalitis A Clinical and Pathological Study - 101 108Document7 pagesSubacute Sclerosing Panencephalitis A Clinical and Pathological Study - 101 108Bhanu PratapNo ratings yet
- Polycystic Kidney - Eight CasesDocument11 pagesPolycystic Kidney - Eight CasesKAINAT ANJUMNo ratings yet
- American Journal of Medical Genetics V PDFDocument2 pagesAmerican Journal of Medical Genetics V PDFBinod KumarNo ratings yet
- International Nosology Heritable Disorders of Connective Tissue, Berlin, 1986Document14 pagesInternational Nosology Heritable Disorders of Connective Tissue, Berlin, 1986Willy AndersonNo ratings yet
- DistoniaDocument5 pagesDistoniaMarlon RojasNo ratings yet
- Sammito1988Document7 pagesSammito1988Aling AyenNo ratings yet
- Becker Muscular Dystrophy Klinefelter's Syndrome: Possible Expression of BMD Within PedigreeDocument4 pagesBecker Muscular Dystrophy Klinefelter's Syndrome: Possible Expression of BMD Within PedigreeYoga LestariNo ratings yet
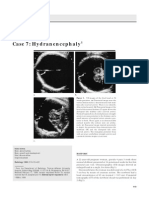
- Case 7: Hydranencephaly: Alfred B. Kurtz, MD Pamela T. Johnson, MDDocument4 pagesCase 7: Hydranencephaly: Alfred B. Kurtz, MD Pamela T. Johnson, MDKenzo Adhi WiranataNo ratings yet
- Avagliano2018 DefectosTuboNeuralDocument13 pagesAvagliano2018 DefectosTuboNeuraliloveshopee9No ratings yet
- Papular Nevus Spilus SyndromeDocument4 pagesPapular Nevus Spilus SyndromePedroNo ratings yet
- Seizure: European Journal of EpilepsyDocument10 pagesSeizure: European Journal of EpilepsyDanilo AssisNo ratings yet
- Cla Eys 1997Document6 pagesCla Eys 1997jahfdfgsdjad asdhsajhajdkNo ratings yet
- Annals of Neurology - 2022 - Gollion - Unilateral Leukoencephalopathy Revealing Cerebral Autosomal Dominant ArteriopathyDocument2 pagesAnnals of Neurology - 2022 - Gollion - Unilateral Leukoencephalopathy Revealing Cerebral Autosomal Dominant Arteriopathyalex mondomobeNo ratings yet
- Girl With Sturge-Weber Syndrome: Case Study 19Document8 pagesGirl With Sturge-Weber Syndrome: Case Study 19CARMEN ELISA NATO SIERRANo ratings yet
- Belfor2006 Clinical and Neuropsychological Features of Corticobasal DegenerationDocument5 pagesBelfor2006 Clinical and Neuropsychological Features of Corticobasal DegenerationEmmanuel Domínguez RosalesNo ratings yet
- Frontotemporal Dementia NeuropathologyDocument14 pagesFrontotemporal Dementia Neuropathologyppico1963No ratings yet
- Autosomal Dominant Inheritance of SpondyloenchondrodysplasiaDocument7 pagesAutosomal Dominant Inheritance of SpondyloenchondrodysplasiaR BNo ratings yet
- Nephrocalcinosis in CretinismDocument4 pagesNephrocalcinosis in CretinismoccuNo ratings yet
- DDH 2 - CompressedDocument105 pagesDDH 2 - CompressedpingryandanNo ratings yet
- Atlas of the Prenatal Mouse BrainFrom EverandAtlas of the Prenatal Mouse BrainUta B. SchambraNo ratings yet
- Kami Export - Jaedyn Griffin - Cellular - RespirationDocument4 pagesKami Export - Jaedyn Griffin - Cellular - RespirationJaedyn GriffinNo ratings yet
- Isolation and Characterization of Cyniclomices GuttulatusDocument7 pagesIsolation and Characterization of Cyniclomices GuttulatusMario Alberto Quintero CaroNo ratings yet
- Bio IA TopsDocument4 pagesBio IA TopsSanjana KumarNo ratings yet
- Biochem 1 Internal Exam QN PaperDocument1 pageBiochem 1 Internal Exam QN PaperVenkatesh TNo ratings yet
- Mechanism of InfectionDocument3 pagesMechanism of Infectionshyamsunder68100% (5)
- Diseases Caused by Bacteria and Viruses.2Document2 pagesDiseases Caused by Bacteria and Viruses.2Melba CuervoNo ratings yet
- Nutrition Guide: National SwineDocument17 pagesNutrition Guide: National SwineEduardo ViolaNo ratings yet
- HERBIVORYDocument39 pagesHERBIVORYDani Mae ManzanilloNo ratings yet
- Manual Fast Prep PDFDocument35 pagesManual Fast Prep PDFNur Insana ImaniarNo ratings yet
- Lab Exercise 5Document2 pagesLab Exercise 5Daks ZabateNo ratings yet
- tmp64C4 TMPDocument12 pagestmp64C4 TMPFrontiersNo ratings yet
- 2019 Horizon Genomic Program Catalogue: HolsteinsDocument7 pages2019 Horizon Genomic Program Catalogue: HolsteinsM Mohsin ChowdhuryNo ratings yet
- 2007 Onwurah, Crude Oils Spills in The Environment Effects and SDocument15 pages2007 Onwurah, Crude Oils Spills in The Environment Effects and SAvanti NurNo ratings yet
- Biochem Lab Report 2Document8 pagesBiochem Lab Report 2Lih XuanNo ratings yet
- Chem 43 - Worksheet3 - NADocument2 pagesChem 43 - Worksheet3 - NAFock StudentNo ratings yet
- MSC BiotechnologyDocument3 pagesMSC Biotechnologyzaki_2611198408No ratings yet
- BIOL 1021 Introductory Biology II: Organismal Biology and Ecology Fall 2019Document23 pagesBIOL 1021 Introductory Biology II: Organismal Biology and Ecology Fall 2019Mariam M. ElgendiNo ratings yet
- BT1001 Biology For Engineers PDFDocument2 pagesBT1001 Biology For Engineers PDFsanthi saranyaNo ratings yet
- Biology S4 SBDocument385 pagesBiology S4 SBbonnealbineNo ratings yet
- NEET-PG 2014 - 2015: Important One Liners For Neet - 5Document22 pagesNEET-PG 2014 - 2015: Important One Liners For Neet - 5adiNo ratings yet
- Gbio Quarter Exam 100 PDFDocument11 pagesGbio Quarter Exam 100 PDFviehazeNo ratings yet
- Science Paper 2 March 2019 STD 10th SSC Maharashtra Board Question PaperDocument4 pagesScience Paper 2 March 2019 STD 10th SSC Maharashtra Board Question PaperSIDDHESH WADEKARNo ratings yet
- SavlonDocument4 pagesSavlonArif MuhammadNo ratings yet
- 2 Regulation Gene Expression BalaccoDocument34 pages2 Regulation Gene Expression Balacco9147Indah WulandariNo ratings yet