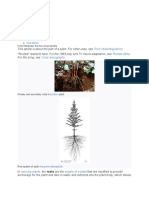
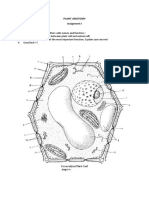
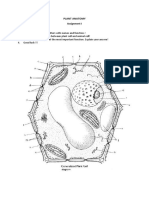
You might also like
- Supporting structures and development in plant leavesDocument10 pagesSupporting structures and development in plant leavesKamilatul KhoirohNo ratings yet
- VeinsDocument5 pagesVeinsMaximos ManiatisNo ratings yet
- 61763885d9fe7e06d3680cbe-1635814495-Leaves - Biology 2eDocument14 pages61763885d9fe7e06d3680cbe-1635814495-Leaves - Biology 2eTresha CosmeñoNo ratings yet
- PLC 7 and 8Document21 pagesPLC 7 and 8rheyukijNo ratings yet
- STONE (Roots&Shoots)Document3 pagesSTONE (Roots&Shoots)Allen KimaniNo ratings yet
- Ephedra: Distribution, Morphological Features and Internal StructuresDocument41 pagesEphedra: Distribution, Morphological Features and Internal StructuresAnkit Jerome FargoseNo ratings yet
- LeavesDocument16 pagesLeavesjasta114No ratings yet
- Plants: The Leaf Adaxial-Abaxial Boundary and Lamina GrowthDocument29 pagesPlants: The Leaf Adaxial-Abaxial Boundary and Lamina Growthlam lamNo ratings yet
- Activity 6 The Root System of PlantsDocument8 pagesActivity 6 The Root System of PlantstscalmaNo ratings yet
- Bidlack, Stern's Introductory Plant Biology, 15e-Highlights LEAFDocument2 pagesBidlack, Stern's Introductory Plant Biology, 15e-Highlights LEAFJasmine SandroNo ratings yet
- A Morphological and Anatomical Observation in Plant LeavesDocument11 pagesA Morphological and Anatomical Observation in Plant LeavesAli SulaimanNo ratings yet
- Ephedra 3Document24 pagesEphedra 3p230552No ratings yet
- Biology - Experiment 7 - LeavesDocument19 pagesBiology - Experiment 7 - LeavesSharlene Tan100% (1)
- Chapter 2. Vegetative Morphology of PlantsDocument23 pagesChapter 2. Vegetative Morphology of PlantsvanisiaNo ratings yet
- Plant Organ Systems: Roots, Stems & LeavesDocument12 pagesPlant Organ Systems: Roots, Stems & LeavesJoebini Aleta RobleteNo ratings yet
- Chapter 4 - Cell and TissueDocument41 pagesChapter 4 - Cell and TissueHanh Hoang100% (1)
- Activity 11 - The Leaf (Instruction Sheet)Document9 pagesActivity 11 - The Leaf (Instruction Sheet)Chris P. BeaconNo ratings yet
- LeafDocument39 pagesLeafDomagoj Butumović100% (1)
- Vascular Tissue DifferentiationDocument9 pagesVascular Tissue DifferentiationC'estMoiNo ratings yet
- Leaf Morphology and AnatomyDocument6 pagesLeaf Morphology and AnatomyVanessaOlgaJ.DagondonNo ratings yet
- Root Meristem, Root Cap, and RootDocument23 pagesRoot Meristem, Root Cap, and RootJulian ManongdoNo ratings yet
- Plant Biology Guide on Organ Systems, Tissues, and StructuresDocument27 pagesPlant Biology Guide on Organ Systems, Tissues, and Structuresashok parmarNo ratings yet
- Chapter 7 - Plant Growth and Options For Reproduction: Plants in ActionDocument58 pagesChapter 7 - Plant Growth and Options For Reproduction: Plants in ActionAgriculture DepartmentNo ratings yet
- Research Paper About LeafDocument10 pagesResearch Paper About LeafGenevieve GanubNo ratings yet
- Monocot Dicot PDFDocument28 pagesMonocot Dicot PDFSunflowerNo ratings yet
- Leaf Anatomy Lab #3: Structures & AdaptationsDocument4 pagesLeaf Anatomy Lab #3: Structures & AdaptationsSafiya JamesNo ratings yet
- Plant Organs Is A Collection of Plant Tissues That Have Specific Functions. Kinds of Plant Organs Are Root, Stem, Leaf and FlowerDocument15 pagesPlant Organs Is A Collection of Plant Tissues That Have Specific Functions. Kinds of Plant Organs Are Root, Stem, Leaf and FlowerkuwukNo ratings yet
- Leaves - Plant Biology: Leaf Arrangements and TypesDocument10 pagesLeaves - Plant Biology: Leaf Arrangements and TypesHarissudin MasrurNo ratings yet
- Stems Lesson 1: Stem FunctionsDocument11 pagesStems Lesson 1: Stem Functionskesyah rubidoNo ratings yet
- Laporan Praktikum Plant Anatomy Anda Physiology Jaringan Pada Daun Monokotil Dan DikotilDocument15 pagesLaporan Praktikum Plant Anatomy Anda Physiology Jaringan Pada Daun Monokotil Dan DikotilMustika RaniNo ratings yet
- PLANTS AND THEIR STRUCTURE - Kuliah 2Document10 pagesPLANTS AND THEIR STRUCTURE - Kuliah 2gahanNo ratings yet
- Growth and Differentiation in PlantsDocument13 pagesGrowth and Differentiation in PlantsApuuli KaraNo ratings yet
- 30.4C: Leaf Structure, Function, and Adaptation: Key PointsDocument3 pages30.4C: Leaf Structure, Function, and Adaptation: Key PointsTresha CosmeñoNo ratings yet
- Plant Anatomy: Plant Body Organization Short Note.: by @tamiyaDocument25 pagesPlant Anatomy: Plant Body Organization Short Note.: by @tamiyaTeamireab DestaNo ratings yet
- Leaf Anatomy: Cells (E), and They Create A Circle Around The Xylem and The Phloem. On TheDocument3 pagesLeaf Anatomy: Cells (E), and They Create A Circle Around The Xylem and The Phloem. On TheANo ratings yet
- How Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersFrom EverandHow Plants Grow - With Information on the Biology of Plant Cells, Roots, Leaves and FlowersNo ratings yet
- Tissue in The Stem Organs of ParenchymaDocument5 pagesTissue in The Stem Organs of Parenchymasandy waskitoNo ratings yet
- Quisao Lab Act 6Document6 pagesQuisao Lab Act 6PaulNo ratings yet
- PlantsDocument14 pagesPlantsGlormina Asprec AvenaNo ratings yet
- FUNARIADocument25 pagesFUNARIAIzzy ChanNo ratings yet
- Primary Plant Body Brainiac ?Document6 pagesPrimary Plant Body Brainiac ?davidoluwadimu28No ratings yet
- WMSU Lab Exercise on Plant TissuesDocument9 pagesWMSU Lab Exercise on Plant TissuesJames Carbonell Dela PeñaNo ratings yet
- Group 2-Excercise 2Document10 pagesGroup 2-Excercise 2James Carbonell Dela PeñaNo ratings yet
- 11 Biology Notes Ch06 Anatomy of Flowering PlantsDocument8 pages11 Biology Notes Ch06 Anatomy of Flowering PlantsPriyanshu BhadanaNo ratings yet
- Paper of Lower Plant TaxonomyDocument35 pagesPaper of Lower Plant TaxonomyMuthiaranifsNo ratings yet
- Lesson Overview: 23.3 StemsDocument29 pagesLesson Overview: 23.3 Stemslayan essamNo ratings yet
- Morphological Features of CycasDocument41 pagesMorphological Features of CycasHamid JoiyaNo ratings yet
- Morphology of MossesDocument11 pagesMorphology of MossesEnrique RodriguezNo ratings yet
- Plant Tissue SystemsDocument2 pagesPlant Tissue SystemsJohn0% (1)
- The main cell types and tissues that make up higher plantsDocument2 pagesThe main cell types and tissues that make up higher plantsJinyuanLowNo ratings yet
- Lesson Prepared Under MHRD Project "National Mission On Education Through ICT"Document39 pagesLesson Prepared Under MHRD Project "National Mission On Education Through ICT"Khalid YousefNo ratings yet
- Reinforcement Plant Growth: Beaconhouse School System Unit: Class: VIIDocument8 pagesReinforcement Plant Growth: Beaconhouse School System Unit: Class: VIIAleena Zahra Naqvi/TCHR/BMBNo ratings yet
- Botany MidtermsDocument16 pagesBotany MidtermsAnisa AzisNo ratings yet
- RootDocument8 pagesRootKristoff Bryan BermacheaNo ratings yet
- Reading Guide Answers PlantsDocument9 pagesReading Guide Answers PlantsNirmal PatelNo ratings yet
- VC molecular control cellular structureDocument17 pagesVC molecular control cellular structureJuan Pablo Matte RisopatronNo ratings yet
- Root Growth and StructureDocument3 pagesRoot Growth and StructureJohn Matthew Jobo100% (1)
- Jaringan Tumbuhan Dalam Bahasa InggrisDocument5 pagesJaringan Tumbuhan Dalam Bahasa InggrisRijal Algifari100% (1)
- StemsDocument2 pagesStemsJela OasinNo ratings yet
- Plant Anatomy: Assignment IDocument1 pagePlant Anatomy: Assignment IKamilatul KhoirohNo ratings yet
- Assignment IDocument1 pageAssignment IKamilatul KhoirohNo ratings yet
- Leaf Base: Occur On The Same Side of The StemDocument1 pageLeaf Base: Occur On The Same Side of The StemKamilatul KhoirohNo ratings yet
- Leaf Surface FeaturesDocument1 pageLeaf Surface FeaturesKamilatul KhoirohNo ratings yet
- 8 Leaf ShapesDocument1 page8 Leaf ShapesKamilatul KhoirohNo ratings yet
- Plant Morphology: Appendages Can Be Distinguish ToDocument1 pagePlant Morphology: Appendages Can Be Distinguish ToKamilatul KhoirohNo ratings yet
- 15 Leaf AttachmentDocument1 page15 Leaf AttachmentKamilatul KhoirohNo ratings yet
- Compound Leaves BreakdownDocument1 pageCompound Leaves BreakdownKamilatul KhoirohNo ratings yet
- 13 Leaf VenationDocument1 page13 Leaf VenationKamilatul KhoirohNo ratings yet
- Specialized Roots: Air, Suck, Insulating & MoreDocument1 pageSpecialized Roots: Air, Suck, Insulating & MoreKamilatul KhoirohNo ratings yet
- LKM Kamilatul Khoiroh - 190210103028Document10 pagesLKM Kamilatul Khoiroh - 190210103028Kamilatul KhoirohNo ratings yet
- Types of Underground Stems and Stem BranchingDocument2 pagesTypes of Underground Stems and Stem BranchingKamilatul KhoirohNo ratings yet
- Types of Palmate and Digitate LeavesDocument1 pageTypes of Palmate and Digitate LeavesKamilatul KhoirohNo ratings yet
- Leaves Are Parts of A Plant or Tree Attached To The Branches or StemsDocument1 pageLeaves Are Parts of A Plant or Tree Attached To The Branches or StemsKamilatul KhoirohNo ratings yet
- 1 Root MorphologyDocument1 page1 Root MorphologyKamilatul KhoirohNo ratings yet
- Practical Work Reports Plant Morphology "The Development of Taproot System and Fibrous Root System"Document21 pagesPractical Work Reports Plant Morphology "The Development of Taproot System and Fibrous Root System"Kamilatul KhoirohNo ratings yet
- Student WorksheetDocument4 pagesStudent WorksheetKamilatul KhoirohNo ratings yet
- Chapter 2. Classification of Ornamental PlantsDocument23 pagesChapter 2. Classification of Ornamental PlantsSaid Abdela100% (4)
- Plan A Vegetable Garden: Seeds or Transplants?Document1 pagePlan A Vegetable Garden: Seeds or Transplants?EthanNo ratings yet
- Calamansi Production Guide: Grow this Acidic Citrus FruitDocument4 pagesCalamansi Production Guide: Grow this Acidic Citrus FruitLando LandoNo ratings yet
- English Homework (Term 1 - 1)Document3 pagesEnglish Homework (Term 1 - 1)TiktoktingzNo ratings yet
- Vol. 2-BoQ, Tech. Spec. & Drawings-Landscape and Irrigation-NCSCMDocument49 pagesVol. 2-BoQ, Tech. Spec. & Drawings-Landscape and Irrigation-NCSCMAbdul Rahuman100% (1)
- t4w zgrTdj3Xe YcDocument10 pagest4w zgrTdj3Xe Ycvink1999No ratings yet
- Mina Rice Cultivation Technology Study in Lebak Swamp LandDocument10 pagesMina Rice Cultivation Technology Study in Lebak Swamp Land30 Raendy ArviansyahNo ratings yet
- Tabebuia AureaDocument3 pagesTabebuia AureaNisamudeen H KklNo ratings yet
- In Vitro Propagation Ludwigia - EgatiDocument12 pagesIn Vitro Propagation Ludwigia - EgatiendangNo ratings yet
- Transform Your Garden Into A ParadiseDocument10 pagesTransform Your Garden Into A ParadiseRenuka ChalikwarNo ratings yet
- Planting and Propagating TreesDocument11 pagesPlanting and Propagating TreesLonlon Lon50% (2)
- Ioannis Therios OlivesDocument426 pagesIoannis Therios OlivesCesar Diaz Barrios100% (3)
- Teakplantation 1Document4 pagesTeakplantation 1Smit kalamkarNo ratings yet
- Seed Starting Information ChartDocument1 pageSeed Starting Information ChartSiraj sirNo ratings yet
- Effect of Gibberellic Acid On Plant Growth (Vigna Radiata)Document5 pagesEffect of Gibberellic Acid On Plant Growth (Vigna Radiata)JackNo ratings yet
- 平行植物 - レオ・レオニDocument214 pages平行植物 - レオ・レオニ西田 編集長100% (1)
- Grassland GrasslandDocument9 pagesGrassland GrasslandMahipal SinghNo ratings yet
- Action Plan-Gulayan Sa PaaralanDocument2 pagesAction Plan-Gulayan Sa PaaralanMarian Oclinaria96% (72)
- Bot - Cot Test - 1Document6 pagesBot - Cot Test - 1Mayur BelsareNo ratings yet
- List of PlantsDocument4 pagesList of PlantsKim KwonNo ratings yet
- Christmas 2014 Catalog OptDocument7 pagesChristmas 2014 Catalog OptEdwin ClemenzNo ratings yet
- Ficus (Fig) Species in Nepal: A Review of Diversity and IndigenousDocument13 pagesFicus (Fig) Species in Nepal: A Review of Diversity and IndigenousamritaryaaligarghNo ratings yet
- Farm Layout and DesignDocument58 pagesFarm Layout and DesignReymond G. Balilu100% (2)
- Types of BarkDocument3 pagesTypes of BarkFrancis Perito100% (1)
- What Plants Need to Grow and ReproduceDocument14 pagesWhat Plants Need to Grow and Reproduceveverica666No ratings yet
- Lysipomia Mitsyae Sp. Nov.Document10 pagesLysipomia Mitsyae Sp. Nov.JEAN SALCEDONo ratings yet
- Amravati Garden Club Amravati Flower Show Category FormatDocument2 pagesAmravati Garden Club Amravati Flower Show Category FormatDr. Dinesh D KhedkarNo ratings yet
- Preserving Physical and Physiological Quality of COCOA SEEDS (Theobroma Cacao L.) DURING STORAGE With The Addition of Organic AcidsDocument18 pagesPreserving Physical and Physiological Quality of COCOA SEEDS (Theobroma Cacao L.) DURING STORAGE With The Addition of Organic AcidsIchaNo ratings yet
- Guide To The Aboriginal Garden Clayton CampusDocument24 pagesGuide To The Aboriginal Garden Clayton CampussinistergripNo ratings yet
- Key Botanical Feature of Safflower PresentationDocument11 pagesKey Botanical Feature of Safflower PresentationSanthosh SanthoshNo ratings yet