You might also like
- Acabado Et Al. 2013 - Fish Larval Flux in Relation To Time and Tide in Taklong National Marine Park, Southern GuimarasDocument8 pagesAcabado Et Al. 2013 - Fish Larval Flux in Relation To Time and Tide in Taklong National Marine Park, Southern GuimarasJose MagallanesNo ratings yet
- Grouper Spawning in CageDocument6 pagesGrouper Spawning in CageHafez MabroukNo ratings yet
- ResearchDocument7 pagesResearchShaera Rose EstropeNo ratings yet
- Milkfish Culture: Alternative Revenue For Mandapam Fisherfolk, Palk Bay, Southeast Coast of IndiaDocument15 pagesMilkfish Culture: Alternative Revenue For Mandapam Fisherfolk, Palk Bay, Southeast Coast of IndiaMike-Dave Madarang CastilloNo ratings yet
- Occurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaDocument10 pagesOccurrence, Distribution and Composition of Marine Mammals in The Bight of Bony, NigeriaRizky RamaNo ratings yet
- 5 StottrupDocument22 pages5 StottrupAdrian Gonzalez GarciaNo ratings yet
- September 2016: See Discussions, Stats, and Author Profiles For This Publication atDocument17 pagesSeptember 2016: See Discussions, Stats, and Author Profiles For This Publication atalpeshmk1No ratings yet
- Growth and Survival of Hatchery Bred GiaDocument15 pagesGrowth and Survival of Hatchery Bred GiaNPCO-PRDP GEF NRMNo ratings yet
- Ecology of WinteringDocument12 pagesEcology of Winteringbiologia007No ratings yet
- Length-Weight Relationship and Fecundity of Liza falcipinnisDocument67 pagesLength-Weight Relationship and Fecundity of Liza falcipinnisHolartoonde_Ho_7736No ratings yet
- 2014 - Notes On Bivalves and Gastropods of PalawanDocument20 pages2014 - Notes On Bivalves and Gastropods of PalawanAireen Diamsim MaltoNo ratings yet
- Seacucumberspawning, LajgsdkjghDocument10 pagesSeacucumberspawning, Lajgsdkjghwawanbio04No ratings yet
- Effect Salinity FW Shrimp 8pDocument8 pagesEffect Salinity FW Shrimp 8pWijaNo ratings yet
- Reef Fishes of Bali, IndonesiaDocument55 pagesReef Fishes of Bali, IndonesiaAchmad DodhyNo ratings yet
- Use of Puerulis Collectors in Lobster Mariculture NJ Quinn BL Kojis 1995Document6 pagesUse of Puerulis Collectors in Lobster Mariculture NJ Quinn BL Kojis 1995Jacque C DiverNo ratings yet
- Pompano SRAC7206Document6 pagesPompano SRAC7206g4nz0No ratings yet
- Anadia Bogotensis-NestingDocument3 pagesAnadia Bogotensis-NestingbiologaemabNo ratings yet
- The Problem and Its BackgroundDocument26 pagesThe Problem and Its BackgroundShainnah CawaNo ratings yet
- MolluscaaaDocument19 pagesMolluscaaaAri OjiNo ratings yet
- Abjna 2 6 897 901Document5 pagesAbjna 2 6 897 901zakiNo ratings yet
- 18607-Article Text-66749-2-10-20180629Document8 pages18607-Article Text-66749-2-10-20180629Aldo OtangNo ratings yet
- Red Claw Densities Study 04Document14 pagesRed Claw Densities Study 04H1H010034No ratings yet
- Aquaculture of Ecteinascidia Turbinata HerdmanDocument6 pagesAquaculture of Ecteinascidia Turbinata HerdmanjlcarballoNo ratings yet
- Enrichment Brachionus.2Document14 pagesEnrichment Brachionus.2Ariez Jack ThepiranhaNo ratings yet
- Biology of Mud CrabDocument8 pagesBiology of Mud CrabMichael jude casaljayNo ratings yet
- Chapter One 1.0 1.1 OystersDocument44 pagesChapter One 1.0 1.1 OystersAkpan EkomNo ratings yet
- Final CHAPTER IIDocument15 pagesFinal CHAPTER IISG DorisAnn Calzado OnceNo ratings yet
- Philippine Aquaculture Overview and HistoryDocument27 pagesPhilippine Aquaculture Overview and Historyvirgil mae taghapNo ratings yet
- REPRODUCTIVE BIOLOGY OF THE ELONGATE SUNSET CLAMDocument55 pagesREPRODUCTIVE BIOLOGY OF THE ELONGATE SUNSET CLAMfenelynNo ratings yet
- Preliminary Checklist of Marine Gastropods and Bivalves in The Kalayaan Island Group Palawan, Western PhilippinesDocument10 pagesPreliminary Checklist of Marine Gastropods and Bivalves in The Kalayaan Island Group Palawan, Western Philippineschoryn modinaNo ratings yet
- Ecological Significance and Commercial Harvesting of Beach-Cast Macro-Algae and Seagrasses in AustraliaDocument16 pagesEcological Significance and Commercial Harvesting of Beach-Cast Macro-Algae and Seagrasses in AustraliaLuane PortelaNo ratings yet
- Distribution and Spatial Structure of Pelagic Fish Schools in Relation To The Nature of The Seabed in The Sicily Straits (Central Mediterranean)Document10 pagesDistribution and Spatial Structure of Pelagic Fish Schools in Relation To The Nature of The Seabed in The Sicily Straits (Central Mediterranean)Ahmad IdrizzNo ratings yet
- Marine Gastropods and Bivalves of Biri, Northern Samar: J.Bio - Innov 8 (3), PP: 319-329, 2019 - ISSN 2277-8330 (Electronic)Document11 pagesMarine Gastropods and Bivalves of Biri, Northern Samar: J.Bio - Innov 8 (3), PP: 319-329, 2019 - ISSN 2277-8330 (Electronic)Claudette RomeroNo ratings yet
- Palma Et Al. 2022. Vet Parasitol 302Document7 pagesPalma Et Al. 2022. Vet Parasitol 302Erlinda LacierdaNo ratings yet
- Carranza 2006 BiolConservDocument6 pagesCarranza 2006 BiolConservtortugamarina100% (2)
- Salt Marsh Ecosystem BudgetDocument33 pagesSalt Marsh Ecosystem Budgetapi-644627281No ratings yet
- Chanos Chanos or Commonly Known As Milkfish. It Is Known in The PhilippinesDocument3 pagesChanos Chanos or Commonly Known As Milkfish. It Is Known in The PhilippinesNathaniel CastasusNo ratings yet
- Infos Tests Analyse DensiteDocument36 pagesInfos Tests Analyse DensiteStanislas MetbaumNo ratings yet
- Larval feeding habits of Maurolicus parvipinnis in Patagonian fjordsDocument12 pagesLarval feeding habits of Maurolicus parvipinnis in Patagonian fjordsjoancoasNo ratings yet
- J Aquaculture 2018 11 042Document29 pagesJ Aquaculture 2018 11 042Miguel García PortaNo ratings yet
- Large-Scale Sandfish (Holothuria Scabra) Aquaculture in Multitrophic Polyculture Ponds in Southern ChinaDocument2 pagesLarge-Scale Sandfish (Holothuria Scabra) Aquaculture in Multitrophic Polyculture Ponds in Southern ChinaKarl KiwisNo ratings yet
- Dermochelys Coriacea (Leatherback Turtle) Listing Advice - Page 1 of 11Document11 pagesDermochelys Coriacea (Leatherback Turtle) Listing Advice - Page 1 of 11ManiNo ratings yet
- 198 Checklist of Fishes Found in The Fresh and Brackish Waters of Negros and Siquijor PhilippinesDocument35 pages198 Checklist of Fishes Found in The Fresh and Brackish Waters of Negros and Siquijor Philippinessyafaiyah dj halidaNo ratings yet
- On The Coast of Pulau Banyak, Indonesia: October 1999Document19 pagesOn The Coast of Pulau Banyak, Indonesia: October 1999Monica KuhonNo ratings yet
- Aqua Farm News Volume 11 (06) November - December 1993Document25 pagesAqua Farm News Volume 11 (06) November - December 1993DENNIS N. MUÑOZNo ratings yet
- Aquaculture in Morocco Growth PerformancDocument12 pagesAquaculture in Morocco Growth PerformancSiham LakhalNo ratings yet
- Articulo Cientifico Q2Document7 pagesArticulo Cientifico Q2Anousha BarriosNo ratings yet
- Turbidity Generation and Biological Impacts of An Exotic Fish PondsDocument10 pagesTurbidity Generation and Biological Impacts of An Exotic Fish PondsHamdi HamzaouiNo ratings yet
- MOLUSKDocument10 pagesMOLUSKElizabeth B DiestroNo ratings yet
- tmp7F30 TMPDocument7 pagestmp7F30 TMPFrontiersNo ratings yet
- Fishery of Commercial Scallops in Asid Gulf, Philippines: Renan U. Bobiles, Victor S. SolimanDocument9 pagesFishery of Commercial Scallops in Asid Gulf, Philippines: Renan U. Bobiles, Victor S. SolimanRed DiggerNo ratings yet
- Naveen-Stromboids Manuscript For+publication+ (1) +Document10 pagesNaveen-Stromboids Manuscript For+publication+ (1) +France Lee ManobanNo ratings yet
- Produção Do PangaDocument8 pagesProdução Do Pangadouglas.eng.worldsNo ratings yet
- Siganus Canaliculatus Park, 1797) in Floating Net With Polyculture SystemDocument10 pagesSiganus Canaliculatus Park, 1797) in Floating Net With Polyculture SystemRafi DarojatNo ratings yet
- Sea Urchin and Sea Cucmber PolycultureDocument13 pagesSea Urchin and Sea Cucmber PolycultureAlentinoNo ratings yet
- Growth and Propagation of Pangasius djambal Catfish in IndonesiaDocument10 pagesGrowth and Propagation of Pangasius djambal Catfish in IndonesiaPascal PascalNo ratings yet
- Effects of Cover and Predator Type on Bufo terrestris Tadpole PredationDocument5 pagesEffects of Cover and Predator Type on Bufo terrestris Tadpole PredationDébora Samira G. NegrãoNo ratings yet
- Darwin's Finches: April 2001Document9 pagesDarwin's Finches: April 2001Rey PinedaNo ratings yet
- 16.2.4 Why Systems Respond Differently To Predator RemovalDocument4 pages16.2.4 Why Systems Respond Differently To Predator RemovalMuhammad AfifudinNo ratings yet
- Whelks to Whales: Coastal Marine Life of the Pacific Northwest, Newly Revised and expanded Third editionFrom EverandWhelks to Whales: Coastal Marine Life of the Pacific Northwest, Newly Revised and expanded Third editionNo ratings yet
- Team-building benefitsDocument3 pagesTeam-building benefitsKarl KiwisNo ratings yet
- Ca 1 - Exercise - CHNDocument2 pagesCa 1 - Exercise - CHNKarl KiwisNo ratings yet
- Pedia Quiz 6Document1 pagePedia Quiz 6Karl KiwisNo ratings yet
- Team-building benefitsDocument3 pagesTeam-building benefitsKarl KiwisNo ratings yet
- Ca 1 Exercise CHN 2Document4 pagesCa 1 Exercise CHN 2Karl KiwisNo ratings yet
- Competency Appraisa1 - ANSWER SHEETDocument1 pageCompetency Appraisa1 - ANSWER SHEETKarl KiwisNo ratings yet
- Summary of Rle KarlDocument12 pagesSummary of Rle KarlKarl KiwisNo ratings yet
- Competency Appraisa1 - ANSWER SHEET 5Document1 pageCompetency Appraisa1 - ANSWER SHEET 5Karl KiwisNo ratings yet
- Competency Appraisal - Pedia - ExerciseDocument2 pagesCompetency Appraisal - Pedia - ExerciseKarl KiwisNo ratings yet
- NavigationDocument8 pagesNavigationKarl Kiwis100% (1)
- Competency Appraisa1 - ANSWER SHEET PREFINALDocument1 pageCompetency Appraisa1 - ANSWER SHEET PREFINALKarl KiwisNo ratings yet
- Competency Appraisal - CHN-3Document2 pagesCompetency Appraisal - CHN-3Karl KiwisNo ratings yet
- Registration: Comprehensive Agrarian Reform ProgramDocument15 pagesRegistration: Comprehensive Agrarian Reform ProgramKarl KiwisNo ratings yet
- Competency Appraisa1 - ANSWER SHEETDocument1 pageCompetency Appraisa1 - ANSWER SHEETKarl KiwisNo ratings yet
- The Earth:: 1. A Prime Meridian Is A Meridian (A Line of Longitude) in A Geographical CoordinateDocument3 pagesThe Earth:: 1. A Prime Meridian Is A Meridian (A Line of Longitude) in A Geographical CoordinateKarl KiwisNo ratings yet
- Extension Education Is An Applied Behavioural Science, The Knowledge of Which Is Applied To BringDocument12 pagesExtension Education Is An Applied Behavioural Science, The Knowledge of Which Is Applied To BringKarl KiwisNo ratings yet
- Second Quarter Lap'S: Microsoft Powerpoint Assessment 1Document3 pagesSecond Quarter Lap'S: Microsoft Powerpoint Assessment 1Karl KiwisNo ratings yet
- AGRARIAN REFORMS DEFINEDDocument20 pagesAGRARIAN REFORMS DEFINEDKarl KiwisNo ratings yet
- The Code of Agrarian Reform (R.A. NO. 3844, AS Amended)Document28 pagesThe Code of Agrarian Reform (R.A. NO. 3844, AS Amended)Karl KiwisNo ratings yet
- Comprehensive Agrarian Reform Program (CARP)Document20 pagesComprehensive Agrarian Reform Program (CARP)Karl KiwisNo ratings yet
- Navigation CHAPTER VDocument16 pagesNavigation CHAPTER VKarl KiwisNo ratings yet
- Diagnostic Procedures OverviewDocument5 pagesDiagnostic Procedures OverviewKarl KiwisNo ratings yet
- Department of Professional Education: First Semester, S.Y. 2018-2019 School of Teacher Education and Liberal Arts (STELA)Document3 pagesDepartment of Professional Education: First Semester, S.Y. 2018-2019 School of Teacher Education and Liberal Arts (STELA)Kristian Karl Bautista Kiw-isNo ratings yet
- BDM19 03 AnonDocument20 pagesBDM19 03 AnonKarl KiwisNo ratings yet
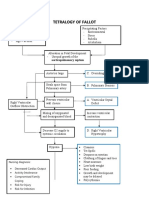
- Tetralogy of Fallot Nursing Diagnosis and ManagementDocument2 pagesTetralogy of Fallot Nursing Diagnosis and ManagementKarl KiwisNo ratings yet
- Diagnostic Procedure PresentationDocument18 pagesDiagnostic Procedure PresentationKarl KiwisNo ratings yet
- 3 Bell Et Al - Aquaculture 273 - 509-519Document12 pages3 Bell Et Al - Aquaculture 273 - 509-519Karl KiwisNo ratings yet
- Potential Nursing Diagnosis: Risk for InfectionDocument2 pagesPotential Nursing Diagnosis: Risk for InfectionKarl KiwisNo ratings yet
- Activity # 3: ISBAR: Kristian Karl B. Kiw-Is BSN Iv-ADocument1 pageActivity # 3: ISBAR: Kristian Karl B. Kiw-Is BSN Iv-AKarl KiwisNo ratings yet
- Community Structure of Echinoderms in Poblacion LiloanDocument24 pagesCommunity Structure of Echinoderms in Poblacion LiloanChristine P. ToledoNo ratings yet
- EchinodermsDocument7 pagesEchinodermsChristian Arm100% (1)
- Echinoderms - Spiny Skin DefendersDocument75 pagesEchinoderms - Spiny Skin DefendersJustyNo ratings yet
- Gia Bianca Diploma in Fish Tech AquacultureDocument27 pagesGia Bianca Diploma in Fish Tech Aquaculturerashelann dalumpinesNo ratings yet
- Echinoderms RevisedDocument8 pagesEchinoderms RevisedPak RisNo ratings yet
- Zoomorphology: Morphology and Function of The Tube Feet of (Echinodermata: Crinoidea)Document13 pagesZoomorphology: Morphology and Function of The Tube Feet of (Echinodermata: Crinoidea)Indra SatriaNo ratings yet
- Zoology Honours Second Semester 2020: Zoocor08: Larva of EchinodermsDocument2 pagesZoology Honours Second Semester 2020: Zoocor08: Larva of EchinodermsRidhi ChoudharyNo ratings yet
- Exam 4 Reading ComprehensionDocument9 pagesExam 4 Reading ComprehensionDewi Sholihah0% (1)
- Chaetognaths Echinoderms and HemichordatesDocument54 pagesChaetognaths Echinoderms and HemichordatesKyrby CabalquintoNo ratings yet
- TOEFL Reading PracticeDocument142 pagesTOEFL Reading PracticeNadia RuizNo ratings yet
- Editan Jan 2017 New-1Document34 pagesEditan Jan 2017 New-1Nisa Nisa100% (1)
- Echinodermata TaxaDocument3 pagesEchinodermata TaxaDave RapaconNo ratings yet
- Sea Cucumber Facts: Diet, Habitat & Health BenefitsDocument18 pagesSea Cucumber Facts: Diet, Habitat & Health BenefitsHoney Belle G. BalderaNo ratings yet
- KIFAYAT Biology NotesDocument135 pagesKIFAYAT Biology NotesAhmad Al FadelNo ratings yet
- Marine BiodiversityDocument178 pagesMarine Biodiversityshanujss100% (1)
- Deichmann 1930 PDFDocument234 pagesDeichmann 1930 PDFAnderson GuerreroNo ratings yet
- Guide to Sandfish Farming for Coastal CommunitiesDocument66 pagesGuide to Sandfish Farming for Coastal CommunitiesDouglas CamposNo ratings yet
- Larva of Echinodermata PDFDocument15 pagesLarva of Echinodermata PDFMilan SarkarNo ratings yet
- Larval Forms in EchinodermataDocument6 pagesLarval Forms in EchinodermatanikhilNo ratings yet
- Phylum Echinodermata: Starfish, Sea Urchin, Sea Brittle, Sea Lilies, Sea Cucumber, Sea FeatherDocument31 pagesPhylum Echinodermata: Starfish, Sea Urchin, Sea Brittle, Sea Lilies, Sea Cucumber, Sea FeatherAmmy SyazannaNo ratings yet
- An Updated Checklist of Echinoderms From Indian WatersDocument68 pagesAn Updated Checklist of Echinoderms From Indian WatersRameshNo ratings yet
- Commercially Important Sea Cucumbers of The WorldDocument223 pagesCommercially Important Sea Cucumbers of The WorldcavrisNo ratings yet
- Echinoderms LectureDocument6 pagesEchinoderms LectureMarjorie Rose TeodosioNo ratings yet
- ĐỀ VIP 5 - PHÁT TRIỂN ĐỀ MINH HỌA THAM KHẢO BGD MÔN ANH NĂM 2024 ( MC2 )_hj3Sj661LeDocument19 pagesĐỀ VIP 5 - PHÁT TRIỂN ĐỀ MINH HỌA THAM KHẢO BGD MÔN ANH NĂM 2024 ( MC2 )_hj3Sj661Letrieuhien0978255565No ratings yet
- S-236546 CompletoDocument361 pagesS-236546 CompletoGustavo MathiasNo ratings yet
- Morfometri, Pemijahan, Dan Indeks Kematangan GonadDocument10 pagesMorfometri, Pemijahan, Dan Indeks Kematangan GonadOki PratamaNo ratings yet
- Deep Sea Creatures of New ZealandDocument319 pagesDeep Sea Creatures of New ZealandJulieta de la HuertaNo ratings yet
- Checklist Nematoda Zootaxa 2011Document88 pagesChecklist Nematoda Zootaxa 2011Ana CarolinaNo ratings yet
- Practical Guide Invertebrates PDFDocument61 pagesPractical Guide Invertebrates PDFMOUSTAFA ALEMAMNo ratings yet
- Sherlyn Teh Yi (IBU1804369) - Individual Written Assignment 2Document21 pagesSherlyn Teh Yi (IBU1804369) - Individual Written Assignment 2Sher LynNo ratings yet