You might also like
- AlzheimersDocument127 pagesAlzheimersshanes100% (1)
- 6 Thinking Hats Detailed Model - UpdatedDocument32 pages6 Thinking Hats Detailed Model - Updatedgeetanshi mittalNo ratings yet
- Perspective Is Therapeutic Plasma Exchange A Viable Option For TreatingDocument11 pagesPerspective Is Therapeutic Plasma Exchange A Viable Option For TreatingRaul Rios RitterNo ratings yet
- Science Grade 7Document8 pagesScience Grade 7Lacus ClyneNo ratings yet
- Historical Background of The SchoolDocument2 pagesHistorical Background of The SchoolJoel Daen50% (2)
- Role of Encapsulated Nano Curcumin in Induced Sh-Sy-5y Cell LineDocument8 pagesRole of Encapsulated Nano Curcumin in Induced Sh-Sy-5y Cell LineInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Poster Template - BiochemistryDocument1 pagePoster Template - Biochemistryraven 37No ratings yet
- DECYSYM - Paper - 2-Arachidonoylglycerol Metabolism in Differntly Modulated by Oligomeric and Fibrillar Confromations...Document13 pagesDECYSYM - Paper - 2-Arachidonoylglycerol Metabolism in Differntly Modulated by Oligomeric and Fibrillar Confromations...LucianoIgnacioMudryjNo ratings yet
- Effect of Naringenin On Aluminum Chloride Induced Alzheimer Disease Power PointDocument19 pagesEffect of Naringenin On Aluminum Chloride Induced Alzheimer Disease Power PointPeter AiyedeNo ratings yet
- Photobiomodulation With Near Infrared Light Helmet in A Pilot Placebo Controlled Clinical Trial in Dementia Patients Testing MemorDocument8 pagesPhotobiomodulation With Near Infrared Light Helmet in A Pilot Placebo Controlled Clinical Trial in Dementia Patients Testing MemorarexixNo ratings yet
- NIH Public Access: Author ManuscriptDocument20 pagesNIH Public Access: Author ManuscriptJosephine D CarandangNo ratings yet
- 1 s2.0 S0969996120304940 MainDocument22 pages1 s2.0 S0969996120304940 MainAmanda MosiniNo ratings yet
- Paper AcrylamideDocument13 pagesPaper AcrylamidekusumawardatiNo ratings yet
- Aad 2020060815151833Document19 pagesAad 2020060815151833George DragosNo ratings yet
- ArticlefrontiersDocument10 pagesArticlefrontiersNadègeKouemouNo ratings yet
- Alzheimer's Disease: A Threat To Mankind: Poorti Pandey, Mritunjai Singh, I. S. GambhirDocument16 pagesAlzheimer's Disease: A Threat To Mankind: Poorti Pandey, Mritunjai Singh, I. S. GambhirWafa BadullaNo ratings yet
- Spinocerebellar Ataxia Type 7 (SCA7) Shows A Cone Rod Dystrophy PhenotypeDocument9 pagesSpinocerebellar Ataxia Type 7 (SCA7) Shows A Cone Rod Dystrophy PhenotypeKarina BudijantoNo ratings yet
- Penelitian Hayati: BerkalaDocument6 pagesPenelitian Hayati: BerkalaRatih AyuNo ratings yet
- endocannabinoide en alzheimerDocument20 pagesendocannabinoide en alzheimersergio serednickiNo ratings yet
- Nihms 841808Document21 pagesNihms 841808Hridhya mpNo ratings yet
- Neuroprotective Effects of Moringa Oleifera Leaf ExtractDocument11 pagesNeuroprotective Effects of Moringa Oleifera Leaf ExtractGREESTYNo ratings yet
- NSAIDs' Role in Alzheimer's Disease via Anti-Amyloidogenic and Anti-Inflammatory EffectsDocument16 pagesNSAIDs' Role in Alzheimer's Disease via Anti-Amyloidogenic and Anti-Inflammatory Effectsdhiraj.2367No ratings yet
- Klein 2006Document13 pagesKlein 2006Dani AlmeidaNo ratings yet
- Amyloid-β in Alzheimer's Disease: Effects on Disease Pathogenesis and Recent Advances in Clinical TrialsDocument8 pagesAmyloid-β in Alzheimer's Disease: Effects on Disease Pathogenesis and Recent Advances in Clinical TrialsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Caldwell 2020 Modeling Neurodegeneration in CaenoDocument28 pagesCaldwell 2020 Modeling Neurodegeneration in CaenoVisa LaserNo ratings yet
- Molecular Tools For The Characterization of Seizure Susceptibility in Genetic Rodent Models of EpilepsyDocument16 pagesMolecular Tools For The Characterization of Seizure Susceptibility in Genetic Rodent Models of EpilepsyJavier HerreroNo ratings yet
- s0047-63742900426-2 Ageing of The Brain Anderton 2002Document7 pagess0047-63742900426-2 Ageing of The Brain Anderton 2002Rifqi Hamdani PasaribuNo ratings yet
- Astrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerDocument15 pagesAstrocytes and Epilepsy: Nihal C. de Lanerolle, Tih-Shih Lee, and Dennis D. SpencerErsin KarataşNo ratings yet
- Kami Export - Chopra, Nanomedicines in The Management of Alzheimer's Disease - Current View and Future ProspectsDocument17 pagesKami Export - Chopra, Nanomedicines in The Management of Alzheimer's Disease - Current View and Future ProspectsДаниал АхатNo ratings yet
- Alcohol Nicotine Proliferation Apoptosis JANG 2002Document5 pagesAlcohol Nicotine Proliferation Apoptosis JANG 2002Andra SilvaNo ratings yet
- Normal aging in rats and pathological aging in human Alzheimer’s disease decrease FAAH activity_ Modulation by cannabinoid agonistsDocument8 pagesNormal aging in rats and pathological aging in human Alzheimer’s disease decrease FAAH activity_ Modulation by cannabinoid agonistssergio serednickiNo ratings yet
- Alzheimer and AcetylcholineDocument14 pagesAlzheimer and Acetylcholinefy fyNo ratings yet
- The Therapeutic Potential of Restoring Gamma Oscillations in Alzheimer's DiseaseDocument9 pagesThe Therapeutic Potential of Restoring Gamma Oscillations in Alzheimer's DiseaseCarlos Hernan Castañeda RuizNo ratings yet
- Microglial Phagocytosis of Neurons in Neurodegeneration, and Its RegulationDocument19 pagesMicroglial Phagocytosis of Neurons in Neurodegeneration, and Its RegulationChristina MountakiNo ratings yet
- Neuroprotective Peptides As Drug Candidates AgainsDocument8 pagesNeuroprotective Peptides As Drug Candidates Againsancuta.lupaescuNo ratings yet
- Andre - Cytomegalic Interneurons - A New Abnormal Cell Type in Severe Pediatric FCDDocument14 pagesAndre - Cytomegalic Interneurons - A New Abnormal Cell Type in Severe Pediatric FCDkudlaceksystemNo ratings yet
- Fulltext Ajcds v4 Id1060Document5 pagesFulltext Ajcds v4 Id1060Malik IbrahimNo ratings yet
- ENEURO.0333-21.2021.fullDocument9 pagesENEURO.0333-21.2021.fullLuciana BenitezNo ratings yet
- Ad ReviewDocument14 pagesAd ReviewKhushNo ratings yet
- Advanced Glycation Endproducts in Peripheral Nerve in Type 2 Diabetes With NeuropathyDocument10 pagesAdvanced Glycation Endproducts in Peripheral Nerve in Type 2 Diabetes With NeuropathyGABYZTNo ratings yet
- Functions and Dysfunctions of Oligodendrocytes inDocument12 pagesFunctions and Dysfunctions of Oligodendrocytes inPriscilla MelindaNo ratings yet
- Effects of Brimonidine On Microglia Morphology in The Transient Retinal Ischemia ModelDocument5 pagesEffects of Brimonidine On Microglia Morphology in The Transient Retinal Ischemia ModelBOHR International Journal of Current Research in Optometry and Ophthalmology (BIJCROO)No ratings yet
- Curcumin Provides Neuroprotection After Spinal Cord Injury: Ó 2009 Elsevier Inc. All Rights ReservedDocument10 pagesCurcumin Provides Neuroprotection After Spinal Cord Injury: Ó 2009 Elsevier Inc. All Rights ReservedanthonyjoelleeNo ratings yet
- NeuralRegenRes1381390-2327675 003847Document6 pagesNeuralRegenRes1381390-2327675 003847JouleAlSBNo ratings yet
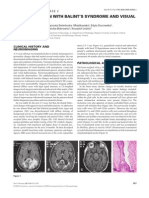
- 31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingDocument4 pages31-Year-Old Man With Balint'S Syndrome and Visual Problems: Clinical History and NeuroimagingwillygopeNo ratings yet
- EEG Analysis Detects Alzheimer's Disease Using Approximate Entropy & Auto Mutual InformationDocument10 pagesEEG Analysis Detects Alzheimer's Disease Using Approximate Entropy & Auto Mutual InformationtourfrikiNo ratings yet
- Structural and Functional Neuroprotection in Glaucoma: Role of Galantamine-Mediated Activation of Muscarinic Acetylcholine ReceptorsDocument11 pagesStructural and Functional Neuroprotection in Glaucoma: Role of Galantamine-Mediated Activation of Muscarinic Acetylcholine ReceptorsFauziah 'pao' Paramita BustamNo ratings yet
- s40120-019-00173-4Document16 pagess40120-019-00173-4Madhu CkNo ratings yet
- Gómez Et Al. 2018Document15 pagesGómez Et Al. 2018Gonzalo Gonzalez AvilaNo ratings yet
- Neuropathologic Changes in ADDocument4 pagesNeuropathologic Changes in ADscientist786No ratings yet
- New Treatments, Tests For Alzheimer'S in PipelineDocument3 pagesNew Treatments, Tests For Alzheimer'S in PipelineChoi Gong JuNo ratings yet
- Early Gene Expression Changes in Spinal Cord From Sod1 Amyotrophic Lateral Sclerosis Animal ModelDocument17 pagesEarly Gene Expression Changes in Spinal Cord From Sod1 Amyotrophic Lateral Sclerosis Animal ModelFabricioNo ratings yet
- LED 660nm Celulas Tronco Do Cerebro Cells-10-01664Document15 pagesLED 660nm Celulas Tronco Do Cerebro Cells-10-01664andre.ramosNo ratings yet
- Syed Ashif Ali: Submitted in The Partial Fulfilment of The Requirements For The Degree ofDocument42 pagesSyed Ashif Ali: Submitted in The Partial Fulfilment of The Requirements For The Degree ofAshif Ali SyedNo ratings yet
- Alzheimer's Disease, CannabinoidsDocument8 pagesAlzheimer's Disease, CannabinoidsPlacido A. Ceballos ChiarucciNo ratings yet
- Articulo STZ IntraventricularDocument16 pagesArticulo STZ Intraventricularp.sepulveda.fiNo ratings yet
- Conduction Block in Acute Motor AxonalDocument12 pagesConduction Block in Acute Motor Axonalgstam69No ratings yet
- BMJ 40 13Document8 pagesBMJ 40 13Alvin JiwonoNo ratings yet
- VIP, Inflammasommes, NMDA, MicrogliaDocument21 pagesVIP, Inflammasommes, NMDA, MicrogliaAhmed NagehNo ratings yet
- Natural Products Targeting Amyloid Beta in Alzheimer's DiseaseDocument17 pagesNatural Products Targeting Amyloid Beta in Alzheimer's Diseaseelver galargaNo ratings yet
- Lee 2021Document17 pagesLee 2021elver galargaNo ratings yet
- Wang, D., & Hazell, A. S. (2010) - Microglial Activation Is A Major Contributor To Neurologic Dysfunction in Thiamine Deficiency PDFDocument6 pagesWang, D., & Hazell, A. S. (2010) - Microglial Activation Is A Major Contributor To Neurologic Dysfunction in Thiamine Deficiency PDFmarej312No ratings yet
- Handbook of Clinical Electrophysiology of VisionFrom EverandHandbook of Clinical Electrophysiology of VisionMinzhong YuNo ratings yet
- Update On Otitis Media - Prevention and Treatment PDFDocument10 pagesUpdate On Otitis Media - Prevention and Treatment PDFangelinefriskaNo ratings yet
- Extracellular Vesicles (Ber)Document13 pagesExtracellular Vesicles (Ber)Farhan Royan PermanahadiNo ratings yet
- The Possible NeuroprotectiveDocument21 pagesThe Possible NeuroprotectiveFarhan Royan PermanahadiNo ratings yet
- A Common Variant of IL-6RDocument27 pagesA Common Variant of IL-6RFarhan Royan PermanahadiNo ratings yet
- Regulation NSC and IL-6 ADDocument13 pagesRegulation NSC and IL-6 ADFarhan Royan PermanahadiNo ratings yet
- MSC-Derived ExosomesDocument22 pagesMSC-Derived ExosomesFarhan Royan PermanahadiNo ratings yet
- Real-Time Monotoring Il-6Document6 pagesReal-Time Monotoring Il-6Farhan Royan PermanahadiNo ratings yet
- MSC-WJ and AD 2 NewDocument12 pagesMSC-WJ and AD 2 NewFarhan Royan PermanahadiNo ratings yet
- MSC Reduce IL-6 NewDocument12 pagesMSC Reduce IL-6 NewFarhan Royan PermanahadiNo ratings yet
- MSC and Il-6Document19 pagesMSC and Il-6Farhan Royan PermanahadiNo ratings yet
- Il-6, Stem Cell, and AdDocument24 pagesIl-6, Stem Cell, and AdFarhan Royan PermanahadiNo ratings yet
- Rapid Review: BackgroundDocument9 pagesRapid Review: BackgroundyusufNo ratings yet
- Potential HNSC and IL-6Document18 pagesPotential HNSC and IL-6Farhan Royan PermanahadiNo ratings yet
- MSC Reduces Il-6 Gene Expression in AdDocument16 pagesMSC Reduces Il-6 Gene Expression in AdFarhan Royan PermanahadiNo ratings yet
- 21.ENG - Oxidative Stress in ADDocument11 pages21.ENG - Oxidative Stress in ADFarhan Royan PermanahadiNo ratings yet
- 25.ENG - Understanding The AD ContinuumDocument10 pages25.ENG - Understanding The AD ContinuumFarhan Royan PermanahadiNo ratings yet
- 38.ENG - The Distribution of Urinary ADDocument10 pages38.ENG - The Distribution of Urinary ADFarhan Royan PermanahadiNo ratings yet
- 30.ENG - The Neuropathological Diagnosis of ADDocument18 pages30.ENG - The Neuropathological Diagnosis of ADFarhan Royan PermanahadiNo ratings yet
- Epidemiology of EncephalitisDocument2 pagesEpidemiology of EncephalitisFarhan Royan PermanahadiNo ratings yet
- 20.ENG - Revisiting The Cholinergic Hypothesis in ADDocument14 pages20.ENG - Revisiting The Cholinergic Hypothesis in ADFarhan Royan PermanahadiNo ratings yet
- 41 Extracellular MSC AdDocument10 pages41 Extracellular MSC AdFarhan Royan PermanahadiNo ratings yet
- Cu Imbalance in ADDocument19 pagesCu Imbalance in ADFarhan Royan PermanahadiNo ratings yet
- AD Patofis, Diagnosis, TerapiDocument11 pagesAD Patofis, Diagnosis, TerapiFarhan Royan PermanahadiNo ratings yet
- XFARDocument14 pagesXFARRIZA SAMPAGANo ratings yet
- A933 Transistor DatasheetDocument5 pagesA933 Transistor Datasheetshorman67134No ratings yet
- Australian Institute For Teaching and School Leadership - AITSLDocument5 pagesAustralian Institute For Teaching and School Leadership - AITSLYu LiNo ratings yet
- Falling Weight Deflectometer (FWD) Projects in IndiaDocument13 pagesFalling Weight Deflectometer (FWD) Projects in IndiaKaran Dave100% (1)
- 2020会社案内(英語版)Document18 pages2020会社案内(英語版)DAC ORGANIZERNo ratings yet
- Dear Boy (Acordes para Cancion)Document4 pagesDear Boy (Acordes para Cancion)Rodolfo GuglielmoNo ratings yet
- LinAlg CompleteDocument331 pagesLinAlg Completes0uizNo ratings yet
- READING U8Document4 pagesREADING U8Như TrầnNo ratings yet
- Capacity Based On Shear Parameters As Per IS 6403: C C C C W Q Q Q Q F y y y yDocument5 pagesCapacity Based On Shear Parameters As Per IS 6403: C C C C W Q Q Q Q F y y y yKishore KumarNo ratings yet
- Williamstown Cannabis Cultivation PlanDocument48 pagesWilliamstown Cannabis Cultivation PlanOltion JaupajNo ratings yet
- Quantitative Data Analysis 2Document10 pagesQuantitative Data Analysis 2Rhona MaandalNo ratings yet
- Staff Nurse HallticketDocument2 pagesStaff Nurse HallticketHiking DiaryNo ratings yet
- Gujrat Newspaper IndustryDocument14 pagesGujrat Newspaper IndustryAlok MahajanNo ratings yet
- Tugas 3vDocument4 pagesTugas 3vRomie SyafitraNo ratings yet
- Food, Family, and FriendshipsDocument256 pagesFood, Family, and FriendshipsBianca PradoNo ratings yet
- Akuntansi Keuangan Lanjutan - Akuntansi Penggabungan UsahaDocument67 pagesAkuntansi Keuangan Lanjutan - Akuntansi Penggabungan UsahachendyNo ratings yet
- Axial Centrifugal FansDocument15 pagesAxial Centrifugal FansAhsan JavedNo ratings yet
- Filed & Entered: SBN 143271 SBN 165797 SBN 259014Document8 pagesFiled & Entered: SBN 143271 SBN 165797 SBN 259014Chapter 11 DocketsNo ratings yet
- V200 User ManualDocument171 pagesV200 User ManualuriahskyNo ratings yet
- SP Manual Addendum For OS 3 - 03Document2 pagesSP Manual Addendum For OS 3 - 03kevmac88No ratings yet
- Duane Grant, A099 743 627 (BIA Sept. 9, 2016)Document12 pagesDuane Grant, A099 743 627 (BIA Sept. 9, 2016)Immigrant & Refugee Appellate Center, LLCNo ratings yet
- Linkstar RCST Installation Guide: Version DDocument56 pagesLinkstar RCST Installation Guide: Version Djorge_chavez01No ratings yet
- Folktalesofkeral 00 MenoDocument124 pagesFolktalesofkeral 00 Menoreena sudhirNo ratings yet
- Orca Share Media1463138182934 PDFDocument167 pagesOrca Share Media1463138182934 PDFJhon Raphael JimenezNo ratings yet
- Collective BargainingDocument18 pagesCollective Bargainingchandni kundel100% (3)
- Strategic Management and Municipal Financial ReportingDocument38 pagesStrategic Management and Municipal Financial ReportingMarius BuysNo ratings yet
- Document Revision TableDocument11 pagesDocument Revision Tableseva1969No ratings yet