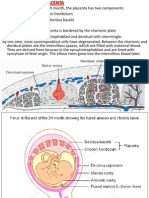
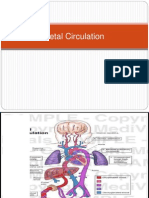
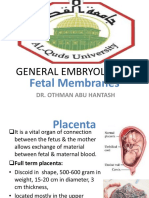
You might also like
- Sin Título 2Document16 pagesSin Título 2Anilu ApNo ratings yet
- Amniotic PhysioDocument3 pagesAmniotic PhysiobjfieroNo ratings yet
- Amniotic Fluid Water Dynamics: M.H. Beall, J.P.H.M. Van Den Wijngaard, M.J.C. Van Gemert, M.G. RossDocument8 pagesAmniotic Fluid Water Dynamics: M.H. Beall, J.P.H.M. Van Den Wijngaard, M.J.C. Van Gemert, M.G. RossCorín AnelNo ratings yet
- Amnioticfluid 120212085244 Phpapp01Document43 pagesAmnioticfluid 120212085244 Phpapp01Jhoanie Sanggoy TauliNo ratings yet
- Pathophysiology of hydrops fetalis and the role of atrial natriuretic peptideDocument25 pagesPathophysiology of hydrops fetalis and the role of atrial natriuretic peptidePandu Nugroho KantaNo ratings yet
- Amniotic Fluid and Its Abnormality: DefinitionDocument7 pagesAmniotic Fluid and Its Abnormality: DefinitionpebripulunganNo ratings yet
- Amniotic Fluid EmbolismDocument34 pagesAmniotic Fluid EmbolismReygie MataNo ratings yet
- OLIGOHIDRAMNIOSDocument20 pagesOLIGOHIDRAMNIOSJorge Luis Sánchez AlarcónNo ratings yet
- Amniotic Fluid As A Vital Sign For Fetal Wellbeing: ReviewDocument9 pagesAmniotic Fluid As A Vital Sign For Fetal Wellbeing: ReviewMeska AmeliaNo ratings yet
- Non-Inmune Hrdrops Fetalis. A Short Review of Etiology and PathophysiologyDocument9 pagesNon-Inmune Hrdrops Fetalis. A Short Review of Etiology and PathophysiologyJuan Manuel RosasNo ratings yet
- Lect12 - Amniotic Fluid ModifiedDocument22 pagesLect12 - Amniotic Fluid Modifiedyelisetti DakshayaniNo ratings yet
- AmnioticDocument7 pagesAmnioticCasey del RosarioNo ratings yet
- Amniotic FluidDocument33 pagesAmniotic FluidElsa Aprillia SujadiNo ratings yet
- Hydra Mi NosDocument8 pagesHydra Mi NosSudha DeviNo ratings yet
- Amniotic FluidDocument55 pagesAmniotic FluidRoselle Joy D. RosalejosNo ratings yet
- Amniotic Fluid: 1 Dr. Mohamed Saad DaoudDocument16 pagesAmniotic Fluid: 1 Dr. Mohamed Saad DaoudVashti SaraswatiNo ratings yet
- Balance de Agua en El Feto y El Recién NacidoDocument5 pagesBalance de Agua en El Feto y El Recién NacidoCarlos ReveloNo ratings yet
- Disorders in Amniotic FluidDocument5 pagesDisorders in Amniotic FluidLatansa DinaNo ratings yet
- Chapter 2Document23 pagesChapter 2Shoroq El BeshbeshyNo ratings yet
- Oligohydramnios - Etiology, Diagnosis, and Management - UpToDateDocument23 pagesOligohydramnios - Etiology, Diagnosis, and Management - UpToDateJUAN FRANCISCO OSORIO PENALOZANo ratings yet
- Amniotic Fluid and ItDocument8 pagesAmniotic Fluid and ItSaman SarKoNo ratings yet
- Physiologic Transition From Intrauterine To Extrauterine LifeDocument12 pagesPhysiologic Transition From Intrauterine To Extrauterine Lifeyhojar PisfilNo ratings yet
- MCN Report Polyhydramnios 1Document24 pagesMCN Report Polyhydramnios 1Vincent CuyucaNo ratings yet
- Physiologic Transition From Intrauterine To Extrauterine Life - UpToDateDocument7 pagesPhysiologic Transition From Intrauterine To Extrauterine Life - UpToDateGuilherme Kazuo OgawaNo ratings yet
- Excess Amniotic Fluid Causes and DiagnosisDocument2 pagesExcess Amniotic Fluid Causes and DiagnosisAde Yonata100% (1)
- Amniocentrices (From The Greek Amnion For "Sac" and KenteDocument5 pagesAmniocentrices (From The Greek Amnion For "Sac" and KenteJohn Paolo Tamayo OrioNo ratings yet
- Pathologies of The Amniotic Fluid Volume: Prepared by DR N. Med. Radosław Blok - I Katedra Gin - Poł. AM WrocławDocument56 pagesPathologies of The Amniotic Fluid Volume: Prepared by DR N. Med. Radosław Blok - I Katedra Gin - Poł. AM WrocławSupreet Singh MalhiNo ratings yet
- 11th Lecture Amnion& Amniotic FluidDocument35 pages11th Lecture Amnion& Amniotic FluidHussein Al Saedi100% (1)
- Oligohydraminos PTDocument8 pagesOligohydraminos PTArpita ArpitaNo ratings yet
- Protocol For Improving Nursing Performance Towards Placental Examination at Labor UnitsDocument11 pagesProtocol For Improving Nursing Performance Towards Placental Examination at Labor Unitsparmar harshilNo ratings yet
- مريم سعد غني GBD OligohydromniospolyhydromniosDocument12 pagesمريم سعد غني GBD OligohydromniospolyhydromniosLubna SaNo ratings yet
- Lecture IV IVDocument37 pagesLecture IV IVMohamad Alsayed AlasaasNo ratings yet
- Amniotic Fluid: Group 5Document29 pagesAmniotic Fluid: Group 5Francis ValdezNo ratings yet
- Dental Management of the Pregnant PatientFrom EverandDental Management of the Pregnant PatientChristos A. SkouterisNo ratings yet
- Amniotic FluidDocument7 pagesAmniotic FluidCherralaine CorderoNo ratings yet
- Physiologic Transition from Fetal to Newborn LifeDocument6 pagesPhysiologic Transition from Fetal to Newborn LifeGina M. TañedoNo ratings yet
- The Amnion and Amniotic FluidDocument7 pagesThe Amnion and Amniotic FluidMerina FirmansahNo ratings yet
- AmnioticfluiddisorderDocument44 pagesAmnioticfluiddisorderNatif BoteNo ratings yet
- Placental Anatomy and Physiology: Ppds Basic 2019Document31 pagesPlacental Anatomy and Physiology: Ppds Basic 2019Andre Putra100% (1)
- Research DCCDocument19 pagesResearch DCCMuhammad ElshamyNo ratings yet
- Polyhydramnios and OligohyramniosDocument35 pagesPolyhydramnios and Oligohyramniosaymanway50% (2)
- Transient Tachypnea of The Newborn: Pediatrics in Review December 2008Document10 pagesTransient Tachypnea of The Newborn: Pediatrics in Review December 2008Haneen Dabbas2019No ratings yet
- Chapter IIDocument11 pagesChapter IIFityan Aulia RahmanNo ratings yet
- Fetal physiologyDocument30 pagesFetal physiologyHassan DalmarNo ratings yet
- Perkembangan Plasenta (Autosaved) .En - IdDocument18 pagesPerkembangan Plasenta (Autosaved) .En - IdFATIMAHNo ratings yet
- 2006 7 E292-E299 William M. Gilbert: Amniotic Fluid DynamicsDocument10 pages2006 7 E292-E299 William M. Gilbert: Amniotic Fluid DynamicsMel MurNo ratings yet
- Wvith: ExchangedDocument10 pagesWvith: ExchangedJessica StephanieNo ratings yet
- Antenatal Physiotherapy Care GuideDocument77 pagesAntenatal Physiotherapy Care Guides_saikalyanNo ratings yet
- Fetal CirculationDocument28 pagesFetal CirculationNelai GoNo ratings yet
- Romzan Alfiriza R (2018) Maryo Juan L.T (2018) Sarah Safrilia (201810401011007)Document22 pagesRomzan Alfiriza R (2018) Maryo Juan L.T (2018) Sarah Safrilia (201810401011007)SarahNo ratings yet
- ch.8 Embryo PDFDocument22 pagesch.8 Embryo PDFMohammad GhannamNo ratings yet
- Hydatidiform Mole: AuthorsDocument7 pagesHydatidiform Mole: AuthorsIdo KurniawanNo ratings yet
- Amniotic Fluid AubfDocument5 pagesAmniotic Fluid AubfVicky ChanNo ratings yet
- Pregnancy Outcome Among Patients With Oligohydramnios and Suggested Plan of ActionDocument11 pagesPregnancy Outcome Among Patients With Oligohydramnios and Suggested Plan of ActionIOSRjournalNo ratings yet
- Dr. Nabeel S. Bondagji, MD, FRCSC: Department of Obstetrics and Gynecology Feto-Maternal UnitDocument24 pagesDr. Nabeel S. Bondagji, MD, FRCSC: Department of Obstetrics and Gynecology Feto-Maternal UnitbeautifulbeastNo ratings yet
- Jurnal Mola HidatidosaDocument8 pagesJurnal Mola HidatidosaDebby SofianaNo ratings yet
- Asfixia NeonatalDocument14 pagesAsfixia NeonatalValeria CuellarNo ratings yet
- Comparative Study of Placental Changes in Mild and Severe Pregnancy HypertensionDocument54 pagesComparative Study of Placental Changes in Mild and Severe Pregnancy HypertensionprasadNo ratings yet
- Amniotic Fluid AbnormalityDocument36 pagesAmniotic Fluid AbnormalityAmanual NuredinNo ratings yet
- Routine Firsttrimester Ultrasound Screening Using A Standardized Anatomical ProtocolDocument15 pagesRoutine Firsttrimester Ultrasound Screening Using A Standardized Anatomical ProtocolYosep SutandarNo ratings yet
- Meta-Analysis of Second-Trimester Markers For Trisomy 21. 2013Document15 pagesMeta-Analysis of Second-Trimester Markers For Trisomy 21. 2013risos7No ratings yet
- Maxillary Gap at 11-13 WeeksDocument5 pagesMaxillary Gap at 11-13 Weekscarlos cobeloNo ratings yet
- PIIS000293781502102XDocument1 pagePIIS000293781502102XYosep SutandarNo ratings yet
- Musa 2015Document15 pagesMusa 2015Jose Luis Morales BautistaNo ratings yet
- Uog 10054Document22 pagesUog 10054Aswin ChristantoNo ratings yet
- IETADocument10 pagesIETAJose Luis Morales BautistaNo ratings yet
- Diagnosis of Fetal Non-Chromosomal Abnormalities On Routine Ultrasound Examination at 11-13 Weeks' GestationDocument10 pagesDiagnosis of Fetal Non-Chromosomal Abnormalities On Routine Ultrasound Examination at 11-13 Weeks' GestationYosep SutandarNo ratings yet
- PDFDocument9 pagesPDFYosep SutandarNo ratings yet
- Ferra RettiDocument51 pagesFerra RettiYosep SutandarNo ratings yet
- Mao2014 Article ImagingOfPelvicLymphNodesDocument13 pagesMao2014 Article ImagingOfPelvicLymphNodesYosep SutandarNo ratings yet
- Pi Is 0002937817310955Document11 pagesPi Is 0002937817310955Yosep SutandarNo ratings yet
- Gestational Trophoblastic NeoplasiaDocument14 pagesGestational Trophoblastic NeoplasiaYosep SutandarNo ratings yet
- Clostridium Perfringens Sepsis Following A Molar Pregnancy: ASE EportDocument2 pagesClostridium Perfringens Sepsis Following A Molar Pregnancy: ASE EportYosep SutandarNo ratings yet
- Murtagh 2010Document9 pagesMurtagh 2010Yosep SutandarNo ratings yet
- PIIS0015028207001161Document8 pagesPIIS0015028207001161Yosep SutandarNo ratings yet
- Changes in The Sexual Function During Pregnancy: Aim. MethodsDocument10 pagesChanges in The Sexual Function During Pregnancy: Aim. MethodsYosep SutandarNo ratings yet
- Effect of Pregnancy On Sexual Function of Couples: AbstractDocument8 pagesEffect of Pregnancy On Sexual Function of Couples: AbstractYosep SutandarNo ratings yet
- Ovarian Reserve Testing: A User's Guide: Expert ReviewsDocument12 pagesOvarian Reserve Testing: A User's Guide: Expert ReviewsYosep SutandarNo ratings yet
- ISUOG - Fetal Heart ExaminationDocument12 pagesISUOG - Fetal Heart ExaminationAzucenaNo ratings yet
- Acog Committee Opinion: Dysmenorrhea and Endometriosis in The AdolescentDocument10 pagesAcog Committee Opinion: Dysmenorrhea and Endometriosis in The AdolescentYosep SutandarNo ratings yet
- Pi Is 0884217517304100Document3 pagesPi Is 0884217517304100Yosep SutandarNo ratings yet
- Practice Bullet In: Emergency ContraceptionDocument11 pagesPractice Bullet In: Emergency ContraceptionYosep SutandarNo ratings yet
- Systematic Reviews To Support Evidence-Based Medicine (2nd Edition)Document216 pagesSystematic Reviews To Support Evidence-Based Medicine (2nd Edition)Yosep Sutandar100% (1)
- Emergency Contraception: Sogc Clinical Practice GuidelineDocument9 pagesEmergency Contraception: Sogc Clinical Practice GuidelineYosep SutandarNo ratings yet
- NSAMR Systematic ReviewDocument8 pagesNSAMR Systematic ReviewYosep SutandarNo ratings yet
- Eng - PDF JsessionidDocument34 pagesEng - PDF JsessionidYosep SutandarNo ratings yet
- FSFI Questionnaire2000Document5 pagesFSFI Questionnaire2000Dewi AsmalindaNo ratings yet
- Enteral NutritionDocument73 pagesEnteral NutritionYosep SutandarNo ratings yet
- SOP of Gram StainDocument5 pagesSOP of Gram Stainzalam55100% (1)
- Grooved Peg BoardDocument2 pagesGrooved Peg BoarddanifgNo ratings yet
- Mental Health Lesson on Identity, Grief, Anxiety, OCD, Schizophrenia, and Eating DisordersDocument2 pagesMental Health Lesson on Identity, Grief, Anxiety, OCD, Schizophrenia, and Eating DisordersKaya S75% (4)
- District Potential Analysis of PilibhitDocument57 pagesDistrict Potential Analysis of PilibhitJasraj SinghNo ratings yet
- 2018 Federal Income Tax Return PDFDocument8 pages2018 Federal Income Tax Return PDFBrandon BachNo ratings yet
- 2) VT Report Test FormatDocument3 pages2) VT Report Test FormatMoin KhanNo ratings yet
- Knime SeventechniquesdatadimreductionDocument266 pagesKnime SeventechniquesdatadimreductionramanadkNo ratings yet
- NURS 366 Exam 1 Study Guide and RubricDocument7 pagesNURS 366 Exam 1 Study Guide and RubriccmpNo ratings yet
- Chapter 18Document10 pagesChapter 18nehallakdawalaNo ratings yet
- DARACLAR 9000HP Silica Brewery Case Study 2016-WebDocument7 pagesDARACLAR 9000HP Silica Brewery Case Study 2016-WebAlia ShabbirNo ratings yet
- Acromegaly: Excess Growth Hormone SecretionDocument4 pagesAcromegaly: Excess Growth Hormone SecretionKavita PathakNo ratings yet
- Points - Discover More RewardsDocument1 pagePoints - Discover More RewardsMustapha El HajjNo ratings yet
- Worksheet#3Document3 pagesWorksheet#3Richard GrayNo ratings yet
- Working at Height PolicyDocument7 pagesWorking at Height PolicyAniekan AkpaidiokNo ratings yet
- Libyan cuisine: Bazin, asida and moreDocument3 pagesLibyan cuisine: Bazin, asida and moreAndy Atehortua CiroNo ratings yet
- EWM Certification Syllabus (9.5) Topic Areas Topic Details, Courses, Books WeightingDocument2 pagesEWM Certification Syllabus (9.5) Topic Areas Topic Details, Courses, Books Weightingguru_3112No ratings yet
- There Came A Gypsy RidingDocument45 pagesThere Came A Gypsy RidingMartin McNelisNo ratings yet
- Cathodic Protection of Subsea Systems: Lessons LearnedDocument9 pagesCathodic Protection of Subsea Systems: Lessons LearnedNguyen Ninh Binh100% (1)
- Plastipak PresentationDocument28 pagesPlastipak Presentationapi-608970005No ratings yet
- Research 1Document7 pagesResearch 1Rhea Mae ObligarNo ratings yet
- Which Statement On Physical Development of Infants and Toddlers Is TRUEDocument9 pagesWhich Statement On Physical Development of Infants and Toddlers Is TRUEKristylle Renz100% (1)
- Beta Analytic Presentation at NNFCC Conference - Linking Bio-Based Materials To Renewable Energy ProductionDocument25 pagesBeta Analytic Presentation at NNFCC Conference - Linking Bio-Based Materials To Renewable Energy ProductionBeta AnalyticNo ratings yet
- Electricity Merit Badge Class 1 2017Document34 pagesElectricity Merit Badge Class 1 2017RoshanNo ratings yet
- VASHISTA AND S1 DEVELOPMENT ONSHORE TERMINALDocument8 pagesVASHISTA AND S1 DEVELOPMENT ONSHORE TERMINALKrm ChariNo ratings yet
- Exercise 5 - Journalizing TransactionsDocument10 pagesExercise 5 - Journalizing TransactionsMichael DiputadoNo ratings yet
- Power Con CorpDocument2 pagesPower Con Corpkhan4luvNo ratings yet
- Electrical Isolation Work PermitDocument2 pagesElectrical Isolation Work PermitGreg GenoveNo ratings yet
- Action On Agricultural. Calculating Agricultural EmissionsDocument27 pagesAction On Agricultural. Calculating Agricultural EmissionsXoel UrriagaNo ratings yet
- The Morning Calm Korea Weekly - Oct. 14, 2005Document26 pagesThe Morning Calm Korea Weekly - Oct. 14, 2005Morning Calm Weekly NewspaperNo ratings yet
- Buller, Paul F. - Schuler, Randall S - Managing Organizations and People - Cases in Management, Organizational Behavior and Human Resource Management-Thomson - South-Western (2006)Document530 pagesBuller, Paul F. - Schuler, Randall S - Managing Organizations and People - Cases in Management, Organizational Behavior and Human Resource Management-Thomson - South-Western (2006)ezekiel capuzNo ratings yet