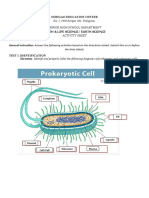
You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (843)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5808)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (346)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Kurbak 2009Document20 pagesKurbak 2009yalocim666No ratings yet
- Jacobson 2004Document8 pagesJacobson 2004yalocim666No ratings yet
- Melbinger 2015Document11 pagesMelbinger 2015yalocim666No ratings yet
- Clarke 2019Document6 pagesClarke 2019yalocim666No ratings yet
- Functioning in Schizophrenia: A Delphi Study Covering The Perspective of PhysiotherapistsDocument9 pagesFunctioning in Schizophrenia: A Delphi Study Covering The Perspective of Physiotherapistsyalocim666No ratings yet
- Tarasidis 1986Document4 pagesTarasidis 1986yalocim666No ratings yet
- Belyuskina 2011Document6 pagesBelyuskina 2011yalocim666No ratings yet
- 10.1007@s00122 001 0795 yDocument9 pages10.1007@s00122 001 0795 yyalocim666No ratings yet
- Yumoto 1997Document15 pagesYumoto 1997yalocim666No ratings yet
- Urits 2021Document16 pagesUrits 2021yalocim666No ratings yet
- Peek 2006Document7 pagesPeek 2006yalocim666No ratings yet
- Lee 2010Document5 pagesLee 2010yalocim666No ratings yet
- Accepted Manuscript: Journal of Physics and Chemistry of SolidsDocument33 pagesAccepted Manuscript: Journal of Physics and Chemistry of Solidsyalocim666No ratings yet
- Pooja 2020Document10 pagesPooja 2020yalocim666No ratings yet
- Ghanbari 2015Document8 pagesGhanbari 2015yalocim666No ratings yet
- Boyd 2021Document10 pagesBoyd 2021yalocim666No ratings yet
- 10 1016@j Jpcs 2019 109217Document52 pages10 1016@j Jpcs 2019 109217yalocim666No ratings yet
- 10 1093@nq@s11-Iv 105 527bDocument1 page10 1093@nq@s11-Iv 105 527byalocim666No ratings yet
- Donahue 1985Document20 pagesDonahue 1985yalocim666No ratings yet
- Jayawickreme 2018Document10 pagesJayawickreme 2018yalocim666No ratings yet
- Zarghamee 1992Document4 pagesZarghamee 1992yalocim666No ratings yet
- 10.1038@s41388 019 1006 0Document13 pages10.1038@s41388 019 1006 0yalocim666No ratings yet
- Zhong 2008Document10 pagesZhong 2008yalocim666No ratings yet
- Mullins 1992Document6 pagesMullins 1992yalocim666No ratings yet
- Akmal 2020Document13 pagesAkmal 2020yalocim666No ratings yet
- Wright 1906Document2 pagesWright 1906yalocim666No ratings yet
- 10 1111@nyas 14211Document23 pages10 1111@nyas 14211yalocim666No ratings yet
- 10 1002@jper 20-0598Document7 pages10 1002@jper 20-0598yalocim666No ratings yet
- Debucquoy 2006Document6 pagesDebucquoy 2006yalocim666No ratings yet
- Longair 2008Document12 pagesLongair 2008yalocim666No ratings yet
- MarcantiaDocument22 pagesMarcantiaRufi RAiNo ratings yet
- Patent Usa MorchellaDocument7 pagesPatent Usa MorchellaViridiana LealNo ratings yet
- CURRICULUM VITAE Pradip Sen4Document6 pagesCURRICULUM VITAE Pradip Sen4Pradip SENNo ratings yet
- Dispersal - of - Seeds - by - Various - Agencies (Mohan) FinalDocument21 pagesDispersal - of - Seeds - by - Various - Agencies (Mohan) FinalKishore Notty100% (1)
- Earth & Life ScienceDocument3 pagesEarth & Life ScienceEHdz D. JameraNo ratings yet
- FloraofMyanmerVII 2021Document12 pagesFloraofMyanmerVII 2021Shine LayNo ratings yet
- 07 - Long-Term On-Farm Participatory Maize Breeding by Stratified Mass Selection Retains Molecular Diversity While Improving Agronomic PerformanceDocument17 pages07 - Long-Term On-Farm Participatory Maize Breeding by Stratified Mass Selection Retains Molecular Diversity While Improving Agronomic PerformanceNemanja NišavićNo ratings yet
- Mendelian Genetics PDFDocument12 pagesMendelian Genetics PDFjhaghjssNo ratings yet
- MODULE 4 in GeneticsDocument6 pagesMODULE 4 in GeneticsChris TianNo ratings yet
- SR Elite, Aiims s60, MPL & LTC Neet Grand Test - 7 Paper (09!07!2022)Document24 pagesSR Elite, Aiims s60, MPL & LTC Neet Grand Test - 7 Paper (09!07!2022)sobhan chinnuNo ratings yet
- DNA Replication Worksheet 2-16-2023 2Document2 pagesDNA Replication Worksheet 2-16-2023 2dfvccfNo ratings yet
- PrintDocument20 pagesPrint4jzbxz64kqNo ratings yet
- Adobe Scan Jun 01, 2022Document18 pagesAdobe Scan Jun 01, 2022xqtfvm8pcsNo ratings yet
- Lucey2014.Distribution of Sponges (Porifera Spongillidae) in Southern Irish Rivers and StreamsDocument13 pagesLucey2014.Distribution of Sponges (Porifera Spongillidae) in Southern Irish Rivers and StreamsAna Paula BertãoNo ratings yet
- Fernández de Palencia, 2008Document31 pagesFernández de Palencia, 200815041168No ratings yet
- Screenshot 2024-01-19 at 5.10.09 PMDocument1 pageScreenshot 2024-01-19 at 5.10.09 PMdhruvrana1133No ratings yet
- 1 January February2023 TBRDocument80 pages1 January February2023 TBRFrank AbrignaniNo ratings yet
- How To Create 3D Plant Cell and Animal Cell Models For Science Class - OwlcationDocument1 pageHow To Create 3D Plant Cell and Animal Cell Models For Science Class - OwlcationMethuli FernandoNo ratings yet
- Biotech Principal and ProcessDocument6 pagesBiotech Principal and ProcessMd Shaukat IqbalNo ratings yet
- Viruses 09 00029 PDFDocument10 pagesViruses 09 00029 PDFJimeno Franscisco de los pozosNo ratings yet
- Aldape 2015Document20 pagesAldape 2015BG BBNo ratings yet
- 18BTB103T - Human Physiology and Health SyllabusDocument3 pages18BTB103T - Human Physiology and Health SyllabusSounak NandyNo ratings yet
- Unit-1 - Nomenclature of EnzymesDocument6 pagesUnit-1 - Nomenclature of Enzymesabhikansh1229No ratings yet
- Board Exams Guide: Blood BankingDocument7 pagesBoard Exams Guide: Blood BankingCindy Mae Flores UtlegNo ratings yet
- 6 Proteins - IntroDocument25 pages6 Proteins - Introemanuel coatesNo ratings yet
- Stem Cell FactorDocument22 pagesStem Cell FactoractivnetNo ratings yet
- Refarat Target Terapi MMDocument57 pagesRefarat Target Terapi MMUmmul HidayahNo ratings yet
- Unit 2: Cell Structure and Function Chapters 6-7Document4 pagesUnit 2: Cell Structure and Function Chapters 6-7api-542216138No ratings yet
- Atlas of The Breeding Birds of Arabia: January 2012Document6 pagesAtlas of The Breeding Birds of Arabia: January 2012Arbi GuetatNo ratings yet
- Aakash Rank Booster Test Series For NEET-2020Document10 pagesAakash Rank Booster Test Series For NEET-2020Siva Shankar SaravananNo ratings yet