You might also like
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Er, Structure & Its FunctionsDocument20 pagesEr, Structure & Its FunctionsTalha AfzalNo ratings yet
- Endoplasmic ReticulumDocument30 pagesEndoplasmic ReticulumMuhammad Junaid Iqbal100% (1)
- Cell STRUCTURE and Function Worksheet. HomeworkDocument5 pagesCell STRUCTURE and Function Worksheet. HomeworkMichael WrightNo ratings yet
- H2 BIO Cell Structure NotesDocument15 pagesH2 BIO Cell Structure NotesEdcademiaNo ratings yet
- ER Role in Protein Synthesis and ModificationDocument56 pagesER Role in Protein Synthesis and Modificationkubuldinho88% (8)
- Biosyntesis Modification, and Cell Secretion 1Document63 pagesBiosyntesis Modification, and Cell Secretion 1Sri HayuniNo ratings yet
- Structure of Endoplasmic Reticulum (ER)Document8 pagesStructure of Endoplasmic Reticulum (ER)Ishita KumariNo ratings yet
- HISTO LEC 2 - The CystoplasmDocument7 pagesHISTO LEC 2 - The CystoplasmjkthmsvtchNo ratings yet
- Mechanism of Intracellular Compartments and Protein Sorting2Document6 pagesMechanism of Intracellular Compartments and Protein Sorting2ahmeddiab2022No ratings yet
- Protein Sorting in A CellDocument26 pagesProtein Sorting in A CellChris AllenNo ratings yet
- Endoplasmic ReticulumDocument12 pagesEndoplasmic ReticulumAzka AltafNo ratings yet
- Zoo Sem2 Endoplasmic ReticulumDocument7 pagesZoo Sem2 Endoplasmic ReticulumAnadi ChauhanNo ratings yet
- Reticulum Endoplasmicum AnsDocument34 pagesReticulum Endoplasmicum AnsNurul MuthiahNo ratings yet
- Biosynthesis, Modification, and Cell Secretion Track: Endoplasmic Reticulum, Ribosome, and Golgi ComplexDocument5 pagesBiosynthesis, Modification, and Cell Secretion Track: Endoplasmic Reticulum, Ribosome, and Golgi ComplexSri HayuniNo ratings yet
- Chapter 4 Cell Review AssignmentDocument8 pagesChapter 4 Cell Review Assignmentsarahmohammed801No ratings yet
- Smooth and Granular Endoplasmic Reticulum RibosomesDocument26 pagesSmooth and Granular Endoplasmic Reticulum RibosomesArfiNo ratings yet
- Lecture 2: Protein Sorting (Endoplasmic Reticulum) : Dr. Diala Abu-Hsasan School of MedicineDocument33 pagesLecture 2: Protein Sorting (Endoplasmic Reticulum) : Dr. Diala Abu-Hsasan School of MedicineAhmad DamatiNo ratings yet
- Cell Biology-Ch12-Part 2-v2Document30 pagesCell Biology-Ch12-Part 2-v2조형윤No ratings yet
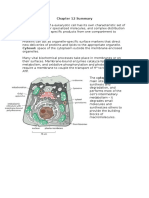
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- Endomembrane System: The Cell's Internal Transport NetworkDocument26 pagesEndomembrane System: The Cell's Internal Transport NetworkPreeti SainiNo ratings yet
- Endoplasmic ReticulumDocument8 pagesEndoplasmic ReticulumShahab Saqib KhanNo ratings yet
- Endomembrane SystemDocument95 pagesEndomembrane SystemSheraz ChaudharyNo ratings yet
- ER Presentation by Abhay AryaDocument17 pagesER Presentation by Abhay AryaAbhay AryaNo ratings yet
- Endomembrane SystemDocument56 pagesEndomembrane SystemSteven Joshua DrizNo ratings yet
- PL4 Cell CompartmentsDocument11 pagesPL4 Cell CompartmentsJake GopitaNo ratings yet
- Endomembrane SystemDocument72 pagesEndomembrane SystemseddadhadilNo ratings yet
- Intracellular Compartment and Protein Sorting RoadmapDocument33 pagesIntracellular Compartment and Protein Sorting RoadmapHaddhi WhibowoNo ratings yet
- MasteringBiology - The Endomembrane SystemDocument11 pagesMasteringBiology - The Endomembrane Systemshiyi33% (3)
- Endoplasmic ReticulumDocument3 pagesEndoplasmic ReticulumHanumat SinghNo ratings yet
- Cell Part 1Document38 pagesCell Part 1Fatih BedirhanNo ratings yet
- 04 Structure To FunctionDocument5 pages04 Structure To Function897291868100% (1)
- Comparison Between Prokaryotic and Eukaryotic Cells Property Prokaryotic Cells Eukaryotic CellsDocument4 pagesComparison Between Prokaryotic and Eukaryotic Cells Property Prokaryotic Cells Eukaryotic CellsKarl Leo Hee YeongNo ratings yet
- Protein TransportDocument13 pagesProtein TransportvmshanesNo ratings yet
- Cell Organelles and FunctionsDocument4 pagesCell Organelles and FunctionsDental LecturesMMQNo ratings yet
- Structure and Functions of Smooth and Rough Endoplasmic ReticulumDocument4 pagesStructure and Functions of Smooth and Rough Endoplasmic Reticulumjahp_13625% (4)
- Biochemical Composition of CellDocument84 pagesBiochemical Composition of Cellmuskaankhan27No ratings yet
- Cellmol-Group12-Final - PPTX 20231023 073441 0000Document29 pagesCellmol-Group12-Final - PPTX 20231023 073441 0000Noronisa PendalidayNo ratings yet
- Zoology CHDocument6 pagesZoology CHAtika SadafNo ratings yet
- Functions of The Rough Endoplasmic Reticulum: Synthesis of Proteins On Membrane-Bound Versus Free RibosomesDocument7 pagesFunctions of The Rough Endoplasmic Reticulum: Synthesis of Proteins On Membrane-Bound Versus Free RibosomesDanie MorenoNo ratings yet
- Endoplasmic RecticulumDocument5 pagesEndoplasmic RecticulumNora ZarahNo ratings yet
- Cell BiologyDocument55 pagesCell Biologyfentaw melkieNo ratings yet
- CELLMOLDocument3 pagesCELLMOLJhon Rey LagosNo ratings yet
- Endoplasmic Reticulum ILLL PDFDocument23 pagesEndoplasmic Reticulum ILLL PDFTIMOTHY JAMESONNo ratings yet
- Cytology 2018Document65 pagesCytology 2018Mohamed ArafaNo ratings yet
- 1583730232049-Endoplasmic ReticulumDocument8 pages1583730232049-Endoplasmic ReticulumGautamiNo ratings yet
- Endoplasmic ReticulumDocument4 pagesEndoplasmic Reticulumnikki_fuentes_1100% (1)
- Assignment Botany 21Document6 pagesAssignment Botany 21Ayesha SadiquaNo ratings yet
- VI CytoplasmDocument68 pagesVI CytoplasmELEONORA MĂRGINEANNo ratings yet
- Cell OrganellesDocument12 pagesCell OrganellesVeronica AlvaradoNo ratings yet
- Biochem LecDocument1 pageBiochem Lecnicolas.princessmiracleb.kldNo ratings yet
- 4, 5 - The Cytoplasm - 2Document58 pages4, 5 - The Cytoplasm - 2Muthana Bani YassenNo ratings yet
- Cell Organelles ExplainedDocument4 pagesCell Organelles ExplainedVeronica AlvaradoNo ratings yet
- Animal Cells IDocument3 pagesAnimal Cells Irandomscribduser2012No ratings yet
- Membranes Part 2 StudentDocument55 pagesMembranes Part 2 StudentBrownyNo ratings yet
- Plant CellDocument8 pagesPlant CellIrish Nicole PacionNo ratings yet
- Partoza - JB, Concept Article - Endomembranesystem - DocxDocument4 pagesPartoza - JB, Concept Article - Endomembranesystem - DocxJenny PartozaNo ratings yet
- BY Arif Mohiddin Mds 1 Year Dept of Oral PathologyDocument40 pagesBY Arif Mohiddin Mds 1 Year Dept of Oral PathologyArif MohiddinNo ratings yet
- 6.1-How Are Cells Studied?: Magnification Resolution Contrast Electron Microscope (EM)Document7 pages6.1-How Are Cells Studied?: Magnification Resolution Contrast Electron Microscope (EM)Sadasivarao GaddeNo ratings yet
- Ana PhyDocument7 pagesAna Physilva, april joy c.No ratings yet
- FRONDA Ethnobiology 2022Document7 pagesFRONDA Ethnobiology 2022Jericho D. FrondaNo ratings yet
- Methods and Principles of Biological SystematicsDocument41 pagesMethods and Principles of Biological SystematicsJericho D. FrondaNo ratings yet
- Cell Biology Midterm Exam ReviewDocument6 pagesCell Biology Midterm Exam ReviewJericho D. FrondaNo ratings yet
- Special Problem Fronda Environmental Quality MonitoringDocument11 pagesSpecial Problem Fronda Environmental Quality MonitoringJericho D. FrondaNo ratings yet
- Chapter VI The Evolution of Plant DiversityDocument36 pagesChapter VI The Evolution of Plant DiversityJericho D. FrondaNo ratings yet
- Fronda - CMB - PPT - Protein TraffickingDocument13 pagesFronda - CMB - PPT - Protein TraffickingJericho D. FrondaNo ratings yet
- Biology Master PDFDocument7 pagesBiology Master PDFRae Krystylynne CodillaNo ratings yet
- NMAT REFRESHER REVIEW: 101616 VERBAL SECTION 1 AND 2Document4 pagesNMAT REFRESHER REVIEW: 101616 VERBAL SECTION 1 AND 2MaverickNo ratings yet
- Repellent Activity of Citrus Oils Against The Cockroaches BlattellaDocument12 pagesRepellent Activity of Citrus Oils Against The Cockroaches BlattellaJericho D. FrondaNo ratings yet
- Repellent Activity of Citrus Oils Against The Cockroaches BlattellaDocument12 pagesRepellent Activity of Citrus Oils Against The Cockroaches BlattellaJericho D. FrondaNo ratings yet
- Microstructures, Mechanical Properties, and Fracture Behaviors of Metal-Injection Molded 17-4PH Stainless SteelDocument7 pagesMicrostructures, Mechanical Properties, and Fracture Behaviors of Metal-Injection Molded 17-4PH Stainless SteelCJPATAGAN100% (1)
- Transition Metals HomeworkDocument17 pagesTransition Metals Homeworkkingman14No ratings yet
- OILFIELD - Inside AdvantageDocument174 pagesOILFIELD - Inside AdvantageGLIM100% (1)
- Waste Water TreatmentDocument28 pagesWaste Water TreatmentRoseNavyaNo ratings yet
- Amity Institute of Pharmacy: B Pharm Ist Semester Pharmaceutical Analysis Pawan Kumar GuptaDocument35 pagesAmity Institute of Pharmacy: B Pharm Ist Semester Pharmaceutical Analysis Pawan Kumar Guptapawan kumar guptaNo ratings yet
- Rate Constant Calculations for Chemical ReactionsDocument11 pagesRate Constant Calculations for Chemical ReactionsUmi LatifahNo ratings yet
- JEE Advanced 2019 Paper-1 ChemistryDocument14 pagesJEE Advanced 2019 Paper-1 ChemistryResonance Eduventures100% (2)
- Normally Closed Solenoid Valves PDFDocument55 pagesNormally Closed Solenoid Valves PDFrahulNo ratings yet
- Guidelines for Swimming Pool ApprovalDocument5 pagesGuidelines for Swimming Pool ApprovalNiel Brian VillarazoNo ratings yet
- C882Document4 pagesC882Jorge Luis Arevalo LopezNo ratings yet
- BS en 00046-2-2016Document28 pagesBS en 00046-2-2016amerNo ratings yet
- Awwa C907-91-Word.Document24 pagesAwwa C907-91-Word.Jhon Gomez CastellanosNo ratings yet
- F3055 14aDocument8 pagesF3055 14asenthilNo ratings yet
- Dry Break CouplingDocument2 pagesDry Break CouplingCookiemonNo ratings yet
- 1 s2.0 S0268005X20312984 MainDocument11 pages1 s2.0 S0268005X20312984 MainAlvaro Matus CNo ratings yet
- MolalityDocument10 pagesMolalityClarisse VasquezNo ratings yet
- CR39W Alloy Properties and ApplicationsDocument2 pagesCR39W Alloy Properties and ApplicationsJean-Noël LerouxNo ratings yet
- Kollidon VA 64Document10 pagesKollidon VA 64Rahma WatiNo ratings yet
- Mineralogy ReviewerDocument12 pagesMineralogy ReviewerMildea Gabuya RabangNo ratings yet
- Remove silt and organics from membranesDocument1 pageRemove silt and organics from membraneseduardoNo ratings yet
- ENG101 Acidizing L N2 - #06 - Acidizing Products Jan05Document25 pagesENG101 Acidizing L N2 - #06 - Acidizing Products Jan05danielmcaeNo ratings yet
- Kalro Attachment ReportDocument43 pagesKalro Attachment ReportLishaEliNo ratings yet
- IGC2 Sample AnswersDocument7 pagesIGC2 Sample Answerspapaji75% (4)
- Expansion JointDocument18 pagesExpansion JointNeha BhatiaNo ratings yet
- Maintenance - Piston'sDocument6 pagesMaintenance - Piston'smecambNo ratings yet
- Smoke Project FinalDocument18 pagesSmoke Project FinalFarhan Akram100% (1)
- CHE1503-101 2016 2 B PDFDocument24 pagesCHE1503-101 2016 2 B PDFsal27adamNo ratings yet
- An Overview On Cellulose-Based Material in Tailoring Bio-Hybrid Nanostructured Photocatalysts For Water Treatment and Renewable Energy ApplicationsDocument26 pagesAn Overview On Cellulose-Based Material in Tailoring Bio-Hybrid Nanostructured Photocatalysts For Water Treatment and Renewable Energy ApplicationsRadu ElenaNo ratings yet
- Lab Report: Shahjalal University of Science & Technology, SylhetDocument6 pagesLab Report: Shahjalal University of Science & Technology, SylhetMd Afif AbrarNo ratings yet
- Resonance (Chemistry) : General Characteristics of ResonanceDocument8 pagesResonance (Chemistry) : General Characteristics of ResonanceMUkesh KumarNo ratings yet