You might also like
- Lab Test Results 7 2015Document3 pagesLab Test Results 7 2015Antzee Dancee0% (1)
- FSM and SIBO 2-3-21 - SlidesDocument81 pagesFSM and SIBO 2-3-21 - SlidesNacer Bechou100% (1)
- Lect 5 Bacterial PathogenicityDocument49 pagesLect 5 Bacterial PathogenicitySuha AbdullahNo ratings yet
- Community Health Nursing Part 1Document64 pagesCommunity Health Nursing Part 1Jim Ryan PauloNo ratings yet
- PH Eur 1433 Herbal DrugsDocument2 pagesPH Eur 1433 Herbal DrugsLuisSanabriaSaavedra0% (1)
- Plants 12 00852 v2Document16 pagesPlants 12 00852 v2Antar OfficialNo ratings yet
- Of in ofDocument9 pagesOf in ofManuel PérezNo ratings yet
- Systemic Acquired ResistanceDocument11 pagesSystemic Acquired Resistanceكركشندي أفندي Karkshndy afndyNo ratings yet
- To Blastand Leaf Blight Diseasesin RiceDocument10 pagesTo Blastand Leaf Blight Diseasesin RicevenkatscribdusernameNo ratings yet
- Involvement of The Rab27 Binding Protein Slac2C/Myrip in Insulin ExocytosisDocument11 pagesInvolvement of The Rab27 Binding Protein Slac2C/Myrip in Insulin ExocytosisJoseGonzalezNo ratings yet
- The Pseudomonas Chlororaphis PCL1391 Sigma Regulator Psra Represses The Production of The Antifungal Metabolite Phenazine-1-Carboxamide PDFDocument10 pagesThe Pseudomonas Chlororaphis PCL1391 Sigma Regulator Psra Represses The Production of The Antifungal Metabolite Phenazine-1-Carboxamide PDFNabil Mahesa FahmiNo ratings yet
- Arabidopsis Lab ReportDocument14 pagesArabidopsis Lab Reportapi-342081300No ratings yet
- Arabidopsis Lab ReportDocument14 pagesArabidopsis Lab Reportapi-342081300No ratings yet
- 1 s2.0 S0960982218310406 MainDocument16 pages1 s2.0 S0960982218310406 MainPavithra MohanNo ratings yet
- TPC 2103700Document14 pagesTPC 2103700gabrielpoulsonNo ratings yet
- Isolation and Characterization of AAP1. A Gene Encoding An Alanine/arginine Aminopeptidase in YeastDocument7 pagesIsolation and Characterization of AAP1. A Gene Encoding An Alanine/arginine Aminopeptidase in Yeasttalha saleemNo ratings yet
- SERP-1-a Secreted Poxviral SerpinDocument4 pagesSERP-1-a Secreted Poxviral SerpinactivnetNo ratings yet
- TMP 7 B46Document13 pagesTMP 7 B46FrontiersNo ratings yet
- Salicylic Acid-Mediated Innate Immunity in Arabidopsis Is Regulated by SIZ1 SUMO E3 LigaseDocument12 pagesSalicylic Acid-Mediated Innate Immunity in Arabidopsis Is Regulated by SIZ1 SUMO E3 LigaseZain BaderNo ratings yet
- 927 FullDocument8 pages927 FullCleaver BrightNo ratings yet
- Frontal Theta Asymmetry Changes While WaDocument89 pagesFrontal Theta Asymmetry Changes While WaLuis Gerardo Ochoa MaldonadoNo ratings yet
- 772 PDFDocument12 pages772 PDFZain BaderNo ratings yet
- Ge 14231021Document11 pagesGe 14231021Diane Fernandez BenesioNo ratings yet
- Plant Virus Interaction - OrelvisDocument66 pagesPlant Virus Interaction - OrelvisJoseph Rafael Aguayo RuizNo ratings yet
- Ras CancerDocument17 pagesRas Cancerniluhputu asrinidewiNo ratings yet
- Mammalian Sterile 20-Like Kinases in Tumor SuppressionDocument3 pagesMammalian Sterile 20-Like Kinases in Tumor Suppressionfranciscrick69No ratings yet
- Plant Cell Reports Volume 26 Issue 8 2007 (Doi 10.1007 - s00299-007-0307-x) Xuelian Zheng Wei Deng Keming Luo Hui Duan Yongqin Chen Ric - The Cauliflower Mosaic Virus (CaMV) 35S Promoter SequencDocument9 pagesPlant Cell Reports Volume 26 Issue 8 2007 (Doi 10.1007 - s00299-007-0307-x) Xuelian Zheng Wei Deng Keming Luo Hui Duan Yongqin Chen Ric - The Cauliflower Mosaic Virus (CaMV) 35S Promoter SequencCarlos Julio Nova LopezNo ratings yet
- Aps 20145Document18 pagesAps 20145Adelia RahmaNo ratings yet
- Differential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsisDocument13 pagesDifferential Gene Expression of ARGININE DECARBOXYLASE ADC1 and ADC2 in ArabidopsismNo ratings yet
- Atrip Britt 2009Document9 pagesAtrip Britt 2009tarasbicotterNo ratings yet
- Identification of RNA Silencing Suppressor Encoded by Wheat Blue Dwarf (WBD) PhytoplasmaDocument24 pagesIdentification of RNA Silencing Suppressor Encoded by Wheat Blue Dwarf (WBD) Phytoplasmasneha vermaNo ratings yet
- Caplen, N. (2001) PDFDocument7 pagesCaplen, N. (2001) PDFLateecka R KulkarniNo ratings yet
- Wound Responsive Cis Element in The 5 Upstream Region of Cucumber Ascorbate Oxidase GeneDocument8 pagesWound Responsive Cis Element in The 5 Upstream Region of Cucumber Ascorbate Oxidase GenenurNo ratings yet
- Genomic Journal 17.8.2018Document36 pagesGenomic Journal 17.8.2018Ana NurulNo ratings yet
- The Plant Journal - 2003 - Sutoh - Two Cis Acting Elements Necessary and Sufficient For Gibberellin Upregulated ProteinaseDocument11 pagesThe Plant Journal - 2003 - Sutoh - Two Cis Acting Elements Necessary and Sufficient For Gibberellin Upregulated ProteinaseElizabeth BenedictaNo ratings yet
- Reporter GenesDocument3 pagesReporter GenesnaNo ratings yet
- RNA Interference: Hannon, Nature 418:244-251Document24 pagesRNA Interference: Hannon, Nature 418:244-251VaishaliSharmaNo ratings yet
- Shauni PosterDocument1 pageShauni Posterapi-268469441No ratings yet
- tmpFE04 TMPDocument5 pagestmpFE04 TMPFrontiersNo ratings yet
- Castañeda 2016Document16 pagesCastañeda 2016papahojaloveNo ratings yet
- Dicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesDocument13 pagesDicot and Monocot Plants Differ in Retinoblastoma-Related Protein SubfamiliesAladár Pettkó-SzandtnerNo ratings yet
- López-Pliego2018 Article TranscriptionalStudyOfTheRsmZDocument11 pagesLópez-Pliego2018 Article TranscriptionalStudyOfTheRsmZpapahojaloveNo ratings yet
- ADAR1 Mutation Causes ZBP1-dependent ImmunopathologyDocument36 pagesADAR1 Mutation Causes ZBP1-dependent ImmunopathologyNidhiDesaiNo ratings yet
- Conserved MYC Transcription Factors Play A Key Role in Jasmonate Signaling Both in Tomato and ArabidopsisDocument16 pagesConserved MYC Transcription Factors Play A Key Role in Jasmonate Signaling Both in Tomato and Arabidopsisrizma.mahaNo ratings yet
- Suppressive Effect of Abscisic Acid On Systemic Acquired Resistance in Tobacco PlantsDocument7 pagesSuppressive Effect of Abscisic Acid On Systemic Acquired Resistance in Tobacco PlantsGustavo HoppeNo ratings yet
- Discusión Art. OmicasDocument6 pagesDiscusión Art. OmicasARIZBETH GUADALUPE VILLANUEVA FLORESNo ratings yet
- NeurodegenerationDocument15 pagesNeurodegenerationDaniela AcostaNo ratings yet
- Systemic Acquired Resistance (SAR)Document38 pagesSystemic Acquired Resistance (SAR)Arulsia ANo ratings yet
- Notes For BIO130, University of TorontoDocument54 pagesNotes For BIO130, University of Torontojasmineopara0% (1)
- Genetic ResearchDocument14 pagesGenetic ResearchDr LipseyNo ratings yet
- A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in PlantsDocument3 pagesA Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plantstanuarora09No ratings yet
- Arabidopsis Decuple Mutant Reveals The Importance of Snrk2 Kinases in Osmotic Stress Responses in VivoDocument6 pagesArabidopsis Decuple Mutant Reveals The Importance of Snrk2 Kinases in Osmotic Stress Responses in VivoJet Lee OlimberioNo ratings yet
- s41598 019 46885 3Document16 pagess41598 019 46885 3SubhasisKarmakarNo ratings yet
- Comprehensive Analysis RalfDocument9 pagesComprehensive Analysis RalfRaymond UnderwoodNo ratings yet
- tmp3E4E TMPDocument4 pagestmp3E4E TMPFrontiersNo ratings yet
- Fimmu 10 01066Document16 pagesFimmu 10 01066julia rossoNo ratings yet
- tmp5C3D TMPDocument10 pagestmp5C3D TMPFrontiersNo ratings yet
- To Study The Interaction Between Auxin and Sugar Signal Transduction Pathway in Model Plant SystemDocument30 pagesTo Study The Interaction Between Auxin and Sugar Signal Transduction Pathway in Model Plant SystemmuhammedjamsheerNo ratings yet
- Journal of Plant Physiology: A B B C DDocument11 pagesJournal of Plant Physiology: A B B C DMirela MihalteanuNo ratings yet
- Nrde 3Document5 pagesNrde 3oqab18No ratings yet
- 016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsDocument25 pages016 Scas Abstracts Listed Alphabetically by Abstractspdf 19 2016 Scas AbstractsdssagNo ratings yet
- Leishmania Amazonensis: Arginase Compartmentalization in The Glycosome Is Important For Parasite InfectivityDocument12 pagesLeishmania Amazonensis: Arginase Compartmentalization in The Glycosome Is Important For Parasite InfectivitynaianaleottiNo ratings yet
- Pnas 202001270Document10 pagesPnas 202001270diego.battiatoNo ratings yet
- Genetics and Biotechnology of BacilliFrom EverandGenetics and Biotechnology of BacilliMark M. ZukowskiNo ratings yet
- Influence of FOX genes on aging and aging-associated diseasesFrom EverandInfluence of FOX genes on aging and aging-associated diseasesNo ratings yet
- Genotype Environment Interaction of Some Traits inDocument14 pagesGenotype Environment Interaction of Some Traits inanicica_866925No ratings yet
- Plant Physiol.-2010-Pitzschke-460-9Document10 pagesPlant Physiol.-2010-Pitzschke-460-9anicica_866925No ratings yet
- JB 06037Document6 pagesJB 06037anicica_866925No ratings yet
- Optimization of Random Amplified Polymorphic Dna Techniques For Use in Genetic Studies of Cuban TriatominaeDocument6 pagesOptimization of Random Amplified Polymorphic Dna Techniques For Use in Genetic Studies of Cuban Triatominaeanicica_866925No ratings yet
- Ijbt 6 (4) 549-552Document4 pagesIjbt 6 (4) 549-552anicica_866925No ratings yet
- Script For Interview (APPLICANT)Document3 pagesScript For Interview (APPLICANT)Hannah Grace Romano ViceralNo ratings yet
- ID4013 - SPW103 - Assessment - 2 - Response Template - 230321 2Document20 pagesID4013 - SPW103 - Assessment - 2 - Response Template - 230321 2SUCHETA DASNo ratings yet
- Shiraishi2014.PDF 02Document6 pagesShiraishi2014.PDF 02noemi leonNo ratings yet
- Listening Sub-Test - Questionpaper: Candidate Details and Photo Will Be Printed HereDocument13 pagesListening Sub-Test - Questionpaper: Candidate Details and Photo Will Be Printed HereNorman NofiantoNo ratings yet
- Disease PreventionDocument14 pagesDisease PreventionJoan InsonNo ratings yet
- Bios AnnualDocument2 pagesBios AnnualGODFREY MULOKOZINo ratings yet
- Journal Article 1 PDFDocument8 pagesJournal Article 1 PDFgerald davidNo ratings yet
- 4H Speech WorksheetDocument2 pages4H Speech WorksheetSTEVENSNo ratings yet
- Cal 21242 UnDocument10 pagesCal 21242 UnLilian Carolina Loreto MachadoNo ratings yet
- Etextbook 978 0134019192 Microbiology With Diseases by TaxonomyDocument62 pagesEtextbook 978 0134019192 Microbiology With Diseases by Taxonomylashawn.fain938100% (49)
- DR Angelo Smith M.D WHPLDocument95 pagesDR Angelo Smith M.D WHPLMuhammad Riaz BhattiNo ratings yet
- HSP Science Form 5Document35 pagesHSP Science Form 5Rafizal ShafieeNo ratings yet
- Interpretation: LPL - PSC Chandigarh Sector 19-D Booth No:-3, Sector:-19-D ChandigarhDocument5 pagesInterpretation: LPL - PSC Chandigarh Sector 19-D Booth No:-3, Sector:-19-D ChandigarhSanNo ratings yet
- Parasitology - Introduction To Medical ParasitologyDocument5 pagesParasitology - Introduction To Medical ParasitologyMarlex SuanNo ratings yet
- Rowen 2018Document7 pagesRowen 2018Regiane AmorimNo ratings yet
- WeeklyBulletin CGSDocument21 pagesWeeklyBulletin CGSJan smithNo ratings yet
- English7 LAS 5 Week 5Document10 pagesEnglish7 LAS 5 Week 5Edmond GalvezNo ratings yet
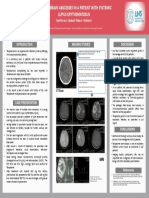
- Discussion Imaging Studies: Multiple Brain Abscesses in A Patient With Systemic Lupus ErythematosusDocument1 pageDiscussion Imaging Studies: Multiple Brain Abscesses in A Patient With Systemic Lupus ErythematosusSolo UpdateNo ratings yet
- MD Pathology Dissertation TopicsDocument6 pagesMD Pathology Dissertation TopicsBuyCustomPaperCanada100% (1)
- Geoff PlattDocument4 pagesGeoff PlattDonnyj1981No ratings yet
- Gamil Sayed Gamil ZeedanDocument48 pagesGamil Sayed Gamil ZeedangamNo ratings yet
- Pyrolysis An Effective Technique For Degradation of COVID 19 Medical WasteDocument20 pagesPyrolysis An Effective Technique For Degradation of COVID 19 Medical WastePietra S.M.No ratings yet
- Young Talk May-June 2011Document5 pagesYoung Talk May-June 2011Straight Talk FoundationNo ratings yet
- Ferris Clinical Advisor 2022 1St Edition Fred F Ferri Full ChapterDocument67 pagesFerris Clinical Advisor 2022 1St Edition Fred F Ferri Full Chaptermichael.walker630100% (15)
- Subjective:: Assessment Diagnosis Planning Implementation Rationale EvaluationDocument2 pagesSubjective:: Assessment Diagnosis Planning Implementation Rationale EvaluationAyra PunzalanNo ratings yet