You might also like
- Nervous Tissue ComponentsDocument15 pagesNervous Tissue ComponentsStar ManavNo ratings yet
- The Nervous SystemDocument5 pagesThe Nervous SystemStephen YorNo ratings yet
- neuroconductivityDocument16 pagesneuroconductivitysandrajoshy06No ratings yet
- Anatomi Fisiologi SarafDocument37 pagesAnatomi Fisiologi SarafUun PrastiyaNo ratings yet
- Nerve Tissue & the Nervous System IntroductionDocument21 pagesNerve Tissue & the Nervous System IntroductionDan UvarovNo ratings yet
- Nervous System Structure and FunctionDocument12 pagesNervous System Structure and FunctionLeon MarkoNo ratings yet
- Nervous TissueDocument25 pagesNervous TissueSumayya KabeerNo ratings yet
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.From EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.No ratings yet
- Cell Body Perikaryon Soma: Neural PlasticityDocument29 pagesCell Body Perikaryon Soma: Neural PlasticitymaNo ratings yet
- Lecture 9 Histology College of Education34351Document6 pagesLecture 9 Histology College of Education34351Zelal Doski100% (1)
- Understanding Nervous TissueDocument30 pagesUnderstanding Nervous TissueOtencianoNo ratings yet
- Nervous System 1 2023Document84 pagesNervous System 1 2023odiodi57No ratings yet
- Nervous Tissue-06.11-2wlDocument57 pagesNervous Tissue-06.11-2wlapi-19641337No ratings yet
- Anatomy of Nervous TissueDocument54 pagesAnatomy of Nervous TissueNand PrakashNo ratings yet
- Structure and Function of NeuronDocument1 pageStructure and Function of NeuronAhsan IqbalNo ratings yet
- 09 Nerve TissueDocument13 pages09 Nerve TissueGovind V PNo ratings yet
- Nervous Tissue: Prof. Dr. Anas Sarwar QureshiDocument12 pagesNervous Tissue: Prof. Dr. Anas Sarwar QureshiShafaqat Ghani Shafaqat GhaniNo ratings yet
- Psy 413 ProjectDocument38 pagesPsy 413 ProjectDumebi AneneNo ratings yet
- NeuronDocument4 pagesNeuroniris danielle barolaNo ratings yet
- Animal Nervous System: Logo HereDocument32 pagesAnimal Nervous System: Logo HereAnne Claudette Capin TeofiloNo ratings yet
- L12 Nerves TissueDocument8 pagesL12 Nerves Tissueعلي اياد شبيب حناشNo ratings yet
- NeuronDocument23 pagesNeuronUnggul YudhaNo ratings yet
- Physiology Handouts (Topic 16-26)Document35 pagesPhysiology Handouts (Topic 16-26)bc200411046No ratings yet
- Neuron and Glial CellsDocument8 pagesNeuron and Glial CellsMurilo Bonet100% (1)
- Nervous Tissue Cells::: Neurons NeurogliaDocument7 pagesNervous Tissue Cells::: Neurons NeurogliaSonakshi KumarNo ratings yet
- THE Nervous System: Biology ProjectDocument8 pagesTHE Nervous System: Biology ProjectAravind JChandranNo ratings yet
- Nervous System: Communications NetworkDocument22 pagesNervous System: Communications NetworkAaron TilAhunNo ratings yet
- 1 Hist Sept 13 ReadingNerve - Tissue2020Document26 pages1 Hist Sept 13 ReadingNerve - Tissue2020n-gorNo ratings yet
- NUR112 Nervous System OverviewDocument30 pagesNUR112 Nervous System OverviewAngel CauilanNo ratings yet
- Group 9 Neuron Structure & FunctionDocument14 pagesGroup 9 Neuron Structure & FunctionMosesNo ratings yet
- Nerve Cells: Structure and FunctionDocument5 pagesNerve Cells: Structure and FunctionSatwant SinghNo ratings yet
- Anatomy and Physiology of The NerveDocument18 pagesAnatomy and Physiology of The NervejudssalangsangNo ratings yet
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- Nervous System (Science Assignment)Document6 pagesNervous System (Science Assignment)Emylya HasnirNo ratings yet
- 1 Cns HistoDocument54 pages1 Cns HistoNaveed AkhterNo ratings yet
- Structure of NeuronsDocument6 pagesStructure of NeuronsMusdiqJavedNo ratings yet
- Anatomical Divisions of The Nervous SystemDocument11 pagesAnatomical Divisions of The Nervous SystemDamianNo ratings yet
- Histology: Neuron Cells Types and StructureDocument7 pagesHistology: Neuron Cells Types and StructureAli HayderNo ratings yet
- Nervous System HistologyDocument4 pagesNervous System HistologyShelly Stephanie Bintoro100% (1)
- Structure of The NeuronDocument7 pagesStructure of The NeuronSmitaNo ratings yet
- Anfis System NeurologiDocument51 pagesAnfis System Neurologianon_822636748No ratings yet
- Neural Control and Co-OrdinationDocument36 pagesNeural Control and Co-Ordinationxanohe8258No ratings yet
- Structure and Functions of Cells of the Nervous SystemDocument48 pagesStructure and Functions of Cells of the Nervous SystemHadia NoorNo ratings yet
- MBBC 2018 Notes 1Document14 pagesMBBC 2018 Notes 1Dimitri KhooNo ratings yet
- Macroglia+Neuron ASG 16-01-2023 PDFDocument9 pagesMacroglia+Neuron ASG 16-01-2023 PDFRajat AgrawalNo ratings yet
- Bilology XII 2014 Ch-17 Nervous CoordinationDocument26 pagesBilology XII 2014 Ch-17 Nervous CoordinationMuhammad UsmanNo ratings yet
- Neuron Structural UnitDocument113 pagesNeuron Structural Unitathena villaNo ratings yet
- Chapter 41 - Neural Signaling (Complete)Document20 pagesChapter 41 - Neural Signaling (Complete)Liana Glorian G. NavarroNo ratings yet
- Nervous TissueDocument101 pagesNervous TissueMitzel SapaloNo ratings yet
- Itzar - Neurohistology PDFDocument186 pagesItzar - Neurohistology PDFahmad aqilNo ratings yet
- Nervous TissueDocument6 pagesNervous TissueAhmed JawdetNo ratings yet
- Acknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsDocument7 pagesAcknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsAndreea LăzăroiuNo ratings yet
- Structure and Function of NeuronsDocument6 pagesStructure and Function of NeuronsAlkykwyn LiteratoNo ratings yet
- NIOT-UnitII-The Nervous System-AnnotationsDocument9 pagesNIOT-UnitII-The Nervous System-Annotationsrt.cseNo ratings yet
- Nervous System BSC 20161Document45 pagesNervous System BSC 20161Arohi ParlikarNo ratings yet
- Summary NeurobiologyDocument42 pagesSummary NeurobiologytjNo ratings yet
- Neural coordination and control in 40 charactersDocument9 pagesNeural coordination and control in 40 charactersYogesh ReddyNo ratings yet
- Table of Content: Serial No Page NoDocument33 pagesTable of Content: Serial No Page NoSID RAJPOOTNo ratings yet
- Neuroanatomy 1Document65 pagesNeuroanatomy 1maNo ratings yet
- ანატომია ზეპირიDocument22 pagesანატომია ზეპირიmaNo ratings yet
- C6, C7, C8, and T1Document20 pagesC6, C7, C8, and T1maNo ratings yet
- UntitledDocument8 pagesUntitledmaNo ratings yet
- Cell Body Perikaryon Soma: Neural PlasticityDocument29 pagesCell Body Perikaryon Soma: Neural PlasticitymaNo ratings yet
- Histology MCQDocument13 pagesHistology MCQHasan Mahmud64% (11)
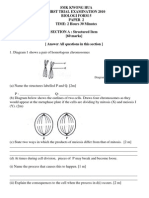
- First Trial Exam - Biologi (SMK Kwong Hua)Document23 pagesFirst Trial Exam - Biologi (SMK Kwong Hua)anakiuNo ratings yet
- Advanced Topics and Applications of AI SyllabusDocument98 pagesAdvanced Topics and Applications of AI Syllabusraghav purohitNo ratings yet
- NEUR 310 SyllabusDocument3 pagesNEUR 310 SyllabusPoudy catNo ratings yet
- Classification Algorithm: Supervised Learning Technique Training DataDocument28 pagesClassification Algorithm: Supervised Learning Technique Training Datamirahaem5No ratings yet
- Nervous SystemDocument2 pagesNervous SystemShanel Aubrey AglibutNo ratings yet
- Consciousness During Cardiac ArrestDocument5 pagesConsciousness During Cardiac ArrestChris SchelinNo ratings yet
- Thalamic Neuron TheoryDocument18 pagesThalamic Neuron TheorybazediNo ratings yet
- Judith Becker - Music and TranceDocument12 pagesJudith Becker - Music and Trancevals100% (1)
- Test Bank For Fundamentals of Human Neuropsychology, 8e Bryan Kolb, Ian Whishaw Test BankDocument7 pagesTest Bank For Fundamentals of Human Neuropsychology, 8e Bryan Kolb, Ian Whishaw Test BankNail BaskoNo ratings yet
- Thyroid HormoneDocument396 pagesThyroid HormoneuiliftengNo ratings yet
- Ana Phisio Lab Report.Document4 pagesAna Phisio Lab Report.Diana Amor100% (1)
- Biology Worksheet G10Document4 pagesBiology Worksheet G10meNo ratings yet
- Heal Your MindDocument96 pagesHeal Your Mindabhilash100% (2)
- NCM 103 Case Analysis 1 (Ms. Perez)Document7 pagesNCM 103 Case Analysis 1 (Ms. Perez)Josefina Isabel GullasNo ratings yet
- Alzheimers Disease and Chediak Hijashi SyndromeDocument47 pagesAlzheimers Disease and Chediak Hijashi Syndromecarmelo aguirreNo ratings yet
- Rstb.2018.0383 Brain Flux ConceptDocument12 pagesRstb.2018.0383 Brain Flux ConceptHua Hidari YangNo ratings yet
- Types of Animal TissueDocument5 pagesTypes of Animal TissueDebiprasad GhoshNo ratings yet
- Biology N5 Past Paper Questions Multicellular Organisms1Document42 pagesBiology N5 Past Paper Questions Multicellular Organisms1Craig MitchellNo ratings yet
- NBSS AIA - Ivan Christian ChannelDocument128 pagesNBSS AIA - Ivan Christian ChannelASTAGINA NAURAHNo ratings yet
- Olatunji AdewaleDocument125 pagesOlatunji AdewaleEbenezer Amoah-KyeiNo ratings yet
- Year 10 Science Progress Assessment: Biology: Paper 1: Biology 1 Higher TierDocument20 pagesYear 10 Science Progress Assessment: Biology: Paper 1: Biology 1 Higher TierYusuf ButtNo ratings yet
- SynapsesDocument46 pagesSynapseshan abuur ahmedNo ratings yet
- Vibrators CT ScreedDocument20 pagesVibrators CT ScreedMuhaiminNo ratings yet
- Determinism, Brain Function and Free Will - Peter G. H. ClarkeDocument18 pagesDeterminism, Brain Function and Free Will - Peter G. H. ClarkeCarlos Augusto VailattiNo ratings yet
- Care For Your BrainDocument265 pagesCare For Your BrainSuresh BalrajNo ratings yet
- Organization of NSDocument20 pagesOrganization of NSnadine azmyNo ratings yet
- Neurosceince Fill in The BlankDocument2 pagesNeurosceince Fill in The Blankjane dwyerNo ratings yet
- Mubarek Aplied Neuroscience PPT Lecture NoteDocument50 pagesMubarek Aplied Neuroscience PPT Lecture NoteAklile TsegaNo ratings yet
- Muscle Physiology and NeurophysiologyDocument68 pagesMuscle Physiology and NeurophysiologyAlan MagpantayNo ratings yet