You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Carlson 2000Document13 pagesCarlson 2000Mamadou DIENENo ratings yet
- The Milan System, From Its Introduction To Its Current Adoption in The Diagnosis of Salivary Gland CytologyDocument9 pagesThe Milan System, From Its Introduction To Its Current Adoption in The Diagnosis of Salivary Gland CytologyMamadou DIENENo ratings yet
- Histopathological Aspects of The Prognostic Factors For Salivary Gland CancersDocument19 pagesHistopathological Aspects of The Prognostic Factors For Salivary Gland CancersMamadou DIENENo ratings yet
- The Epigenesis of Salivary Glands CarcinomaDocument31 pagesThe Epigenesis of Salivary Glands CarcinomaMamadou DIENENo ratings yet
- Salivary Gland FNA Diagnostics in A Real-LifeDocument11 pagesSalivary Gland FNA Diagnostics in A Real-LifeMamadou DIENENo ratings yet
- Complication SubmandibulectomieDocument5 pagesComplication SubmandibulectomieMamadou DIENENo ratings yet
- Altérations de L'histologie de La Glande Salivaire Vieillissante Et Corrélation Avec L'inflammation GlandulaireDocument22 pagesAltérations de L'histologie de La Glande Salivaire Vieillissante Et Corrélation Avec L'inflammation GlandulaireMamadou DIENENo ratings yet
- Effets Pronostiques de La Chirurgie Ou de La Radiothérapie Sur Le Carcinome Adénoïde Kystique de La Tête Et Du CouDocument17 pagesEffets Pronostiques de La Chirurgie Ou de La Radiothérapie Sur Le Carcinome Adénoïde Kystique de La Tête Et Du CouMamadou DIENENo ratings yet
- L'innervation Parasympathique Dans La Glande SalivaireDocument14 pagesL'innervation Parasympathique Dans La Glande SalivaireMamadou DIENENo ratings yet
- Interactions Entre Les Nerfs en Développement Et Les Glandes SalivairesDocument8 pagesInteractions Entre Les Nerfs en Développement Et Les Glandes SalivairesMamadou DIENENo ratings yet
- Prise en Charge Des Tumeurs Des Glandes Salivaires Royaume-Uni Lignes Directrices Nationales MultidisciplinairesDocument8 pagesPrise en Charge Des Tumeurs Des Glandes Salivaires Royaume-Uni Lignes Directrices Nationales MultidisciplinairesMamadou DIENENo ratings yet
- Oral Histology Salivary GlandsDocument10 pagesOral Histology Salivary GlandsMamadou DIENENo ratings yet
- Ghafoor SalivaryGlandsDocument6 pagesGhafoor SalivaryGlandsMamadou DIENENo ratings yet
- Medical English1 1Document4 pagesMedical English1 1Mamadou DIENENo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Histomorphology of The Pancreas in MynaDocument6 pagesHistomorphology of The Pancreas in MynaIndian Journal of Veterinary and Animal Sciences RNo ratings yet
- Histology of GitDocument31 pagesHistology of GitLucky LuckyNo ratings yet
- Histo NotesDocument19 pagesHisto Notesanas ishaqNo ratings yet
- Pancreas+salivary GlandsDocument4 pagesPancreas+salivary GlandsAllen BurdowskiNo ratings yet
- HistologyDocument109 pagesHistologyMEHMET ALİ KIRBIYIKNo ratings yet
- Group 3 Sjogrens SyndromeDocument9 pagesGroup 3 Sjogrens SyndromeFrancine LerioNo ratings yet
- Histology: First Semester A.Y. 2021 - 2022Document22 pagesHistology: First Semester A.Y. 2021 - 2022Fea Kristine PacquiaoNo ratings yet
- 16: Salivary Gland Dev. and Structure (Complete)Document7 pages16: Salivary Gland Dev. and Structure (Complete)NYUCD17No ratings yet
- SALIVADocument68 pagesSALIVANimmi RenjithNo ratings yet
- Endocrine System - MedDocument124 pagesEndocrine System - Medrediet shimekachNo ratings yet
- Oral Histology & Embryology-FikreDocument240 pagesOral Histology & Embryology-FikreHeran A AlhadiNo ratings yet
- 10 Digestive System: C.T. Reyes, DVM, MSDocument26 pages10 Digestive System: C.T. Reyes, DVM, MSKent Solis VicenteNo ratings yet
- Lab 04 Glandular EpitheliumDocument2 pagesLab 04 Glandular EpitheliumSarwar JafarNo ratings yet
- Salivary GlandsDocument14 pagesSalivary Glandsandre gidion laseNo ratings yet
- Salivary GlandsDocument97 pagesSalivary GlandsSwati George100% (2)
- Anatomy Histology of The Oral Cavity2 3Document31 pagesAnatomy Histology of The Oral Cavity2 3drsondosNo ratings yet
- Gall Bladder & Pancreas HistologyDocument48 pagesGall Bladder & Pancreas HistologyGood BoyNo ratings yet
- Digestive HistologyDocument11 pagesDigestive HistologyFahd Abdullah Al-refai100% (2)
- Histology Drawings PDFDocument19 pagesHistology Drawings PDFVishwanath Sinduvadi100% (1)
- Histology of GitDocument56 pagesHistology of GitDoc On CallNo ratings yet
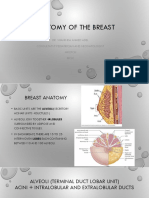
- Anatomy of The Breast1Document13 pagesAnatomy of The Breast1Shahinda Ahmed AdelNo ratings yet
- Histology AssignmentDocument4 pagesHistology AssignmentNico LokoNo ratings yet
- Glandular EpitheliumDocument10 pagesGlandular EpitheliumAbby Claire SomeraNo ratings yet
- Annals of Anatomy: Hitomi Ono Minagi, Yu Usami, Manabu Sakai, Takayoshi SakaiDocument7 pagesAnnals of Anatomy: Hitomi Ono Minagi, Yu Usami, Manabu Sakai, Takayoshi SakaiMamadou DIENENo ratings yet
- Gastrointestinal Tract 1Document14 pagesGastrointestinal Tract 1Jezzah Mae CañeteNo ratings yet