You might also like
- A-level Biology Revision: Cheeky Revision ShortcutsFrom EverandA-level Biology Revision: Cheeky Revision ShortcutsRating: 5 out of 5 stars5/5 (5)
- Lecture 3 Notes Cellular Basis of LifeDocument11 pagesLecture 3 Notes Cellular Basis of Lifeochienganthony001No ratings yet
- Chapter 3 CellDocument94 pagesChapter 3 Cellmohamedshilleh7No ratings yet
- CSIR Life Science Sample Cell BiologyDocument17 pagesCSIR Life Science Sample Cell Biologygamer zoneNo ratings yet
- ANSWEREEDocument3 pagesANSWEREERaxilla MaxillaNo ratings yet
- Struct-Function Cell OrgaDocument30 pagesStruct-Function Cell OrgaSiaw Sing OngNo ratings yet
- Study Exercise Biology 10-12 97.03Document7 pagesStudy Exercise Biology 10-12 97.03Fabiola Vania FeliciaNo ratings yet
- Chapter 5Document20 pagesChapter 5skywalkerNo ratings yet
- Biochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TextDocument60 pagesBiochemistry & Genetics: II SHS 109: Resource Person: DR Tanveer Akbar Reference TexttNo ratings yet
- Kuliah MembranDocument61 pagesKuliah Membrannikmah nuur rochmahNo ratings yet
- 16.plasma membr-WPS OfficeDocument4 pages16.plasma membr-WPS OfficeMohamed RizwanNo ratings yet
- Functional and Structural Properties of Natural BiomembranesDocument7 pagesFunctional and Structural Properties of Natural BiomembranesAhsan RazaNo ratings yet
- Cell Biology: Boundary LipidDocument18 pagesCell Biology: Boundary LipidVittal Athul RNo ratings yet
- 4-Biological Membrane and Transport 21-22Document21 pages4-Biological Membrane and Transport 21-22sarahwassel2015No ratings yet
- L2 Membranes 14 15 STDocument62 pagesL2 Membranes 14 15 STLara WeberNo ratings yet
- Chapter 7Document4 pagesChapter 7Astrii LyNo ratings yet
- Pale Child with Splenomegaly and Hemolytic AnemiaDocument49 pagesPale Child with Splenomegaly and Hemolytic AnemiaAwais RehmanNo ratings yet
- 5 - Composition and Functions of Cell Membrane Transport Systems LectureDocument32 pages5 - Composition and Functions of Cell Membrane Transport Systems Lecturegodara28pNo ratings yet
- Cell Membrane Structure & FunctionsDocument2 pagesCell Membrane Structure & FunctionsKim Antonette50% (2)
- The Cellular Level of OrganizationDocument7 pagesThe Cellular Level of OrganizationPearl MirandaNo ratings yet
- Cell Membrane (Plasma Membrane)Document6 pagesCell Membrane (Plasma Membrane)Rica NorcioNo ratings yet
- Intracellular TransportDocument66 pagesIntracellular Transportalvitakhoridatul100% (1)
- Cell Membranes (20025)Document52 pagesCell Membranes (20025)MUHAMMAD USAMANo ratings yet
- The Cell MembraneDocument36 pagesThe Cell Membraneanulorance98No ratings yet
- Chapter 5 Lesson 1-Cell Structures Involved in TransportDocument8 pagesChapter 5 Lesson 1-Cell Structures Involved in TransportDaichi KatashiNo ratings yet
- Cell Biology HandoutsDocument37 pagesCell Biology HandoutsIanDiel ParagosoNo ratings yet
- Lesson 2 Cell Membrane and Lesson 3 HomeostasisDocument14 pagesLesson 2 Cell Membrane and Lesson 3 HomeostasisAj MirandaNo ratings yet
- 2 The Cell - Structure and Function 228964Document17 pages2 The Cell - Structure and Function 228964Musharaf RehmanNo ratings yet
- Cell MembraneDocument5 pagesCell MembranejmcvicenteNo ratings yet
- Cell TransportDocument2 pagesCell TransportAshleyNo ratings yet
- BS PUBLIC HEALTH BIOLOGY LECTURE MEMBRANE STRUCTUREDocument8 pagesBS PUBLIC HEALTH BIOLOGY LECTURE MEMBRANE STRUCTUREJohn Michael OsorioNo ratings yet
- Membrane Structure and Function: Lecture OutlineDocument11 pagesMembrane Structure and Function: Lecture OutlineJohn Westly S. SabueroNo ratings yet
- 9gkneylgi4kueakdv8q9gkney50iscvv SolutionDocument2 pages9gkneylgi4kueakdv8q9gkney50iscvv SolutionArya MohajerNo ratings yet
- Plasma Membrane The Big PictureDocument4 pagesPlasma Membrane The Big PictureNicolas Gomez CombellasNo ratings yet
- Cell Functions & Organelles in 40 CharactersDocument14 pagesCell Functions & Organelles in 40 Charactersamber tariqNo ratings yet
- Cellular-Organization Part1Document7 pagesCellular-Organization Part1Nina IlaganNo ratings yet
- 1 2 3 4 5 6 MergedDocument90 pages1 2 3 4 5 6 MergedNAYEEMA JAMEEL ANUVANo ratings yet
- C9e Answers Active Reading 07Document7 pagesC9e Answers Active Reading 07Daniel DHizzy Hizgilov83% (18)
- Cell Theory and Biochemical Aspects of Cell MembraneDocument39 pagesCell Theory and Biochemical Aspects of Cell MembraneHaroon BadarNo ratings yet
- Cell Membrane Transport ActivityDocument7 pagesCell Membrane Transport ActivityoctavianojanineNo ratings yet
- General Biology 1 Reviewer 1Document5 pagesGeneral Biology 1 Reviewer 1Shane Patricia CalmaNo ratings yet
- Genbio Plasma Membrane ReviewerDocument3 pagesGenbio Plasma Membrane ReviewerEleah MelchorNo ratings yet
- Cell Membrane and Cell WallDocument4 pagesCell Membrane and Cell WallViral DaftaryNo ratings yet
- Medical Biology 1: Cytology and Cell Organelles/TITLEDocument48 pagesMedical Biology 1: Cytology and Cell Organelles/TITLEFady FadyNo ratings yet
- Chapter7 - Membrane Structure and FunctionDocument13 pagesChapter7 - Membrane Structure and FunctionKrizziane Ivy CuajotorNo ratings yet
- Physio 1 Day MainDocument302 pagesPhysio 1 Day Mainanup raoNo ratings yet
- Simple Notes - Paper I PhysiologyDocument147 pagesSimple Notes - Paper I PhysiologyArrya DSNo ratings yet
- NRS1102: Anatomy and Physiology (Lecture) 1 Semester 2020-2021 Ejfmania DMD, RN, MANDocument14 pagesNRS1102: Anatomy and Physiology (Lecture) 1 Semester 2020-2021 Ejfmania DMD, RN, MANEllen Mynelle MabulacNo ratings yet
- 002 Cell Membrane PhysiologyDocument12 pages002 Cell Membrane PhysiologyAlfred BajarNo ratings yet
- Cell MembraneDocument17 pagesCell MembraneEloisa MadrilenoNo ratings yet
- BIO 315. NotesDocument42 pagesBIO 315. NotesKevin KipropNo ratings yet
- Online Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreDocument16 pagesOnline Practice Tests, Live Classes, Tutoring, Study Guides Q&A, Premium Content and MoreYoAmoNYCNo ratings yet
- Lec 1Document39 pagesLec 1Hussain AhmedNo ratings yet
- Concise Histology Gartner, Leslie PDFDocument478 pagesConcise Histology Gartner, Leslie PDFLexsshNo ratings yet
- Mader/Biology, 11/e - Chapter Outline: 5.1 Plasma Membrane Structure and FunctionDocument4 pagesMader/Biology, 11/e - Chapter Outline: 5.1 Plasma Membrane Structure and Functionapi-455371000No ratings yet
- Cell and Molecular BiologyDocument2 pagesCell and Molecular BiologyAttaullah36No ratings yet
- Unit 2 ReviewerDocument4 pagesUnit 2 Revieweraaleah moscaNo ratings yet
- Plasma Membrane DeutschDocument5 pagesPlasma Membrane DeutschSafeer SefiNo ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesFrom EverandCellular and Molecular Mechanisms of Inflammation: Vascular Adhesion MoleculesCharles G. CochraneNo ratings yet
- People v. Lagarto, 326 SCRA 693 (2000)Document43 pagesPeople v. Lagarto, 326 SCRA 693 (2000)GioNo ratings yet
- 20ME901 Automobile Engineering Unit 3Document74 pages20ME901 Automobile Engineering Unit 36044 sriramNo ratings yet
- Soal Kelas 4 IIDocument5 pagesSoal Kelas 4 IIes tougeNo ratings yet
- Eagle Test ReportDocument25 pagesEagle Test ReportMuhammad FahadNo ratings yet
- Solutions: Spheres, Cones and CylindersDocument13 pagesSolutions: Spheres, Cones and CylindersKeri-ann MillarNo ratings yet
- 41 PDFsam Redis CookbookDocument5 pages41 PDFsam Redis CookbookHữu Hưởng NguyễnNo ratings yet
- Inventory Storage and Retrieval System PatentDocument15 pagesInventory Storage and Retrieval System PatentdevanasokaNo ratings yet
- ProjectDocument86 pagesProjectrajuNo ratings yet
- Act 1&2 and SAQ No - LawDocument4 pagesAct 1&2 and SAQ No - LawBududut BurnikNo ratings yet
- Astm D 664 - 07Document8 pagesAstm D 664 - 07Alfonso MartínezNo ratings yet
- Essene Moses Ten CommandmentsDocument4 pagesEssene Moses Ten Commandmentsjeryon91No ratings yet
- Oracle Apps Quality ModuleDocument17 pagesOracle Apps Quality ModuleSantOsh100% (2)
- Visualization BenchmarkingDocument15 pagesVisualization BenchmarkingRanjith S100% (1)
- Digital Water Monitoring and Turbidity Quality System Using MicrocontrollerDocument8 pagesDigital Water Monitoring and Turbidity Quality System Using MicrocontrollerIrin DwiNo ratings yet
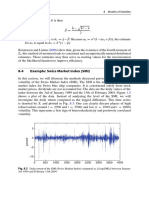
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- spl400 Stereo Power Amplifier ManualDocument4 pagesspl400 Stereo Power Amplifier ManualRichter SiegfriedNo ratings yet
- C-Dot Max-XlDocument39 pagesC-Dot Max-XlGourav Roy100% (3)
- One - Pager - SOGEVAC SV 320 BDocument2 pagesOne - Pager - SOGEVAC SV 320 BEOLOS COMPRESSORS LTDNo ratings yet
- 2nd QUARTER EXAMINATION IN P. E 2019-2020Document3 pages2nd QUARTER EXAMINATION IN P. E 2019-2020Lyzl Mahinay Ejercito MontealtoNo ratings yet
- Class 7thDocument24 pagesClass 7thPriyaNo ratings yet
- Oas Community College: Republic of The Philippines Commission On Higher Education Oas, AlbayDocument22 pagesOas Community College: Republic of The Philippines Commission On Higher Education Oas, AlbayJaycel NepalNo ratings yet
- Proposed Panel Antenna: Globe Telecom ProprietaryDocument2 pagesProposed Panel Antenna: Globe Telecom ProprietaryJason QuibanNo ratings yet
- Bhakti Trader Ram Pal JiDocument232 pagesBhakti Trader Ram Pal JiplancosterNo ratings yet
- Characteristics and Guidelines of PublicspaceDocument3 pagesCharacteristics and Guidelines of PublicspaceJanani SurenderNo ratings yet
- Service Parts List: 54-26-0005 2551-20 M12™ FUEL™ SURGE™ 1/4" Hex Hydraulic Driver K42ADocument2 pagesService Parts List: 54-26-0005 2551-20 M12™ FUEL™ SURGE™ 1/4" Hex Hydraulic Driver K42AAmjad AlQasrawi100% (1)
- INVENTORY MANAGEMENT TechniquesDocument24 pagesINVENTORY MANAGEMENT TechniquesWessal100% (1)
- Starting A Business Candle Making 2009Document2 pagesStarting A Business Candle Making 2009Carlo Fabros Junio100% (4)
- Fehniger Steve-Pump VibrationDocument35 pagesFehniger Steve-Pump VibrationSheikh Shoaib100% (1)
- ClientsDocument7 pagesClientsLiane PanahacNo ratings yet
- Aerospace Opportunities in SwitzerlandDocument5 pagesAerospace Opportunities in SwitzerlandBojana DekicNo ratings yet