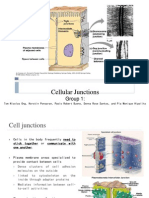
You might also like
- Mammalian Histology AssignmentDocument9 pagesMammalian Histology AssignmentSana Sultana100% (1)
- Cellular JunctionsDocument25 pagesCellular JunctionsDiana Grace Pile0% (1)
- Epithelium Surface SpecializationsDocument41 pagesEpithelium Surface SpecializationsMitzel SapaloNo ratings yet
- NPTEL-Tissue Engineering Cell ContactDocument6 pagesNPTEL-Tissue Engineering Cell ContactHarini BalasubramanianNo ratings yet
- Integrating Cells Into Tissues: OutlineDocument71 pagesIntegrating Cells Into Tissues: OutlineEvelin Cornejo FundoraNo ratings yet
- Cell JunctionsDocument1 pageCell JunctionsJoshua Mari De OcampoNo ratings yet
- Basal Lamina: Foundation of Epithelial-Connective Tissue InterfaceDocument8 pagesBasal Lamina: Foundation of Epithelial-Connective Tissue InterfaceMegan LewisNo ratings yet
- Cell Junctions Form TissuesDocument5 pagesCell Junctions Form TissuesDan ChoiNo ratings yet
- Carcinoma Invasion and Metastasis A Role For EpithelialDocument5 pagesCarcinoma Invasion and Metastasis A Role For Epithelialfranciscrick69No ratings yet
- EG' Histology (5th Ed.) Bookmarks and SearchableDocument325 pagesEG' Histology (5th Ed.) Bookmarks and SearchableCHRISTOPHER JOHN MANINGASNo ratings yet
- Cell Junction - KBK RahmanDocument12 pagesCell Junction - KBK RahmanSukhrian MuhdaNo ratings yet
- Cell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemDocument70 pagesCell-Cell Adhesion and Cell Junction: Submitted by Ashish Palodkar Msc. Biotechnology 1 SemGovinda BiswasNo ratings yet
- Cell JunctionsDocument29 pagesCell JunctionsArjun DahalNo ratings yet
- Cell Junctions, Cell-Cell AdhesionsDocument36 pagesCell Junctions, Cell-Cell AdhesionsRizky Nur IndahNo ratings yet
- Bab 1 2 Biomembran PDFDocument84 pagesBab 1 2 Biomembran PDFMuh JufriNo ratings yet
- Cell Junctions, Gap Adhesions, and ExtracellularDocument58 pagesCell Junctions, Gap Adhesions, and ExtracellularFabian Christine MaeNo ratings yet
- Human Anatomy 4th Edition Saladin Solutions ManualDocument25 pagesHuman Anatomy 4th Edition Saladin Solutions ManualLeslieBuchanannjfy100% (51)
- Cold Spring Harb Perspect Biol-2015-PoberDocument12 pagesCold Spring Harb Perspect Biol-2015-PoberpamelaNo ratings yet
- EEU2020 Adfinex HDocument23 pagesEEU2020 Adfinex HZainab Jamal SiddiquiNo ratings yet
- Type of Cell Junctions 1Document12 pagesType of Cell Junctions 1shahrul mukminNo ratings yet
- Full Download Human Anatomy 4th Edition Saladin Solutions ManualDocument35 pagesFull Download Human Anatomy 4th Edition Saladin Solutions Manualvocality.duettoqw603100% (36)
- Cell JunctionsDocument56 pagesCell JunctionsDrGagandeep Singh100% (3)
- Basic Cell Theory - StructureDocument32 pagesBasic Cell Theory - StructureRuby SamarNo ratings yet
- JI UNIT 2 Complete Notes - 7-8Document2 pagesJI UNIT 2 Complete Notes - 7-8Shubham PawadeNo ratings yet
- Cell-Cell Interfaces As Specialized Compartments Directing Cell FunctionDocument15 pagesCell-Cell Interfaces As Specialized Compartments Directing Cell FunctionDaoud IssaNo ratings yet
- Understanding the Dynamic CytoskeletonDocument66 pagesUnderstanding the Dynamic CytoskeletonChristian Santiago100% (1)
- Cell Structure: Prepared by Dr. Sana AslamDocument41 pagesCell Structure: Prepared by Dr. Sana Aslamsajjad Ali balouch100% (1)
- BBS1 HS K4 Cell Junction Intercellular CommunicationDocument26 pagesBBS1 HS K4 Cell Junction Intercellular CommunicationVania IlmiNo ratings yet
- Bloc 8 La Cel Lula I L'entornDocument7 pagesBloc 8 La Cel Lula I L'entornLaia Baus RevueltaNo ratings yet
- Adhesion y Migración Celular 2021-1 - CompressedDocument44 pagesAdhesion y Migración Celular 2021-1 - CompressedNiyuhNo ratings yet
- Cell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetDocument16 pagesCell-Cell Junctions: Bhavtosh A. Kikani, PHD, Icar-NetB KikaniNo ratings yet
- Cellular Nanomechanics 2Document27 pagesCellular Nanomechanics 2Ahmed MotasimNo ratings yet
- MorphogenesisDocument25 pagesMorphogenesisDrAbhilasha SharmaNo ratings yet
- Cho2016 Article RolesOfCross-MembraneTransportDocument13 pagesCho2016 Article RolesOfCross-MembraneTransportTaurus PearsonNo ratings yet
- Gap-Junction-Mediated Cell-To-Cell Communication: ReviewDocument11 pagesGap-Junction-Mediated Cell-To-Cell Communication: Reviewdornaz69No ratings yet
- nihms-1923722Document41 pagesnihms-1923722Phạm Trần Minh NhựtNo ratings yet
- B GENERAL BIOLOGY I Q1M3.1 Learner-Copy Final LayoutDocument10 pagesB GENERAL BIOLOGY I Q1M3.1 Learner-Copy Final LayoutLiennar NaikhNo ratings yet
- Assignment Anatomi HakimDocument2 pagesAssignment Anatomi HakimIkmal HakimNo ratings yet
- Histology ReviewerDocument7 pagesHistology ReviewerJulius FrondaNo ratings yet
- Histology (Epithelial Tissue)Document23 pagesHistology (Epithelial Tissue)FazilaNo ratings yet
- MSC Botnay Pre Pap1 bl1 PDFDocument74 pagesMSC Botnay Pre Pap1 bl1 PDFShahid pinNo ratings yet
- Cell JunctionsDocument18 pagesCell Junctionsrevathikarri065No ratings yet
- Cell Junction: 3 Types Intercellular Junctions in AnimalsDocument7 pagesCell Junction: 3 Types Intercellular Junctions in AnimalsNada hasanNo ratings yet
- Cell Junctions: Specialized Cell Junctions Occur at Points of Cell-Cell andDocument8 pagesCell Junctions: Specialized Cell Junctions Occur at Points of Cell-Cell andRahul SharmaNo ratings yet
- PhysiologyDocument205 pagesPhysiologyppdzssx7ghNo ratings yet
- Yannai2018 - Journal of Cell Molecular MedicineDocument6 pagesYannai2018 - Journal of Cell Molecular MedicineolalaulaNo ratings yet
- General Histology Tissue Types and FunctionsDocument133 pagesGeneral Histology Tissue Types and FunctionsHenok GirmaNo ratings yet
- Cellular JunctionsDocument11 pagesCellular JunctionsAhmed Zubair IrshadNo ratings yet
- CELL CYTOSKELETON, CELL JUNCTIONSDocument46 pagesCELL CYTOSKELETON, CELL JUNCTIONSAhmed khanNo ratings yet
- Cells: Nucleus (Eukaryotes) or Nucleoid (Bacteria)Document33 pagesCells: Nucleus (Eukaryotes) or Nucleoid (Bacteria)Leandro GalzeranoNo ratings yet
- Cell CommunicationDocument27 pagesCell Communicationapi-292966101No ratings yet
- Intercellular Junctions in Animals and PlantsDocument22 pagesIntercellular Junctions in Animals and PlantsBrillian AlfiNo ratings yet
- Unit 15 Cell Junction DhanDocument18 pagesUnit 15 Cell Junction DhanSarah PavuNo ratings yet
- Epithelium IDocument33 pagesEpithelium IUmerNo ratings yet
- Cell JunctionsDocument40 pagesCell JunctionsAhmad MursyidNo ratings yet
- Junctions Between CellsDocument2 pagesJunctions Between CellsAljon FresnidoNo ratings yet
- MOOC_Cellular_Regeneration_articleDocument5 pagesMOOC_Cellular_Regeneration_articleahmatogreNo ratings yet
- Complex Networks Orchestrate Epithelial-Mesenchymal Transitions (Σημειωμένο)Document12 pagesComplex Networks Orchestrate Epithelial-Mesenchymal Transitions (Σημειωμένο)panna1No ratings yet
- Parts of The Cell WallDocument28 pagesParts of The Cell WallJOSHUA DICHOSONo ratings yet
- Journeys B2 Teacher's Resource Pack: Grammar ThemesDocument2 pagesJourneys B2 Teacher's Resource Pack: Grammar ThemesMiriam SanchezNo ratings yet
- Pricelist PT SCB Maret 2023Document11 pagesPricelist PT SCB Maret 2023PT RUKUN CAHAYA ABADINo ratings yet
- GasGas TXT Racing 2014 Owner's ManualDocument60 pagesGasGas TXT Racing 2014 Owner's Manualstone chouNo ratings yet
- Liquichek Specialty Immunoassay Control Levels LTA, 1, 2 and 3Document6 pagesLiquichek Specialty Immunoassay Control Levels LTA, 1, 2 and 3clinicalbiochemistryNo ratings yet
- Analyzing Potential Audit Client LakesideDocument22 pagesAnalyzing Potential Audit Client LakesideLet it be100% (1)
- Tools for Theft InvestigationDocument15 pagesTools for Theft InvestigationMalagant EscuderoNo ratings yet
- B.sc. Microbiology 1Document114 pagesB.sc. Microbiology 1nasitha princeNo ratings yet
- Stigmatization of Feminism: Gender Studies As "Gender Ideology" in Right-Wing Populist Political Discourse in HungaryDocument55 pagesStigmatization of Feminism: Gender Studies As "Gender Ideology" in Right-Wing Populist Political Discourse in HungaryAnubhav SinghNo ratings yet
- IT Industry in India: Indian Education SystemDocument9 pagesIT Industry in India: Indian Education SystemPradeep BommitiNo ratings yet
- Device Test ModeDocument14 pagesDevice Test ModeNay SoeNo ratings yet
- Compassionate Sciences - RedactedDocument184 pagesCompassionate Sciences - RedactedNew Jersey marijuana documentsNo ratings yet
- Data Center Cooling Solutions That Lower Costs With High Energy SavingsDocument5 pagesData Center Cooling Solutions That Lower Costs With High Energy Savingskhamsone pengmanivongNo ratings yet
- Heliopolis Language Modern School Academic Year (2015-2016) Model Exam (9) 1st Prep (Unit 3) Aim High (A-Level)Document2 pagesHeliopolis Language Modern School Academic Year (2015-2016) Model Exam (9) 1st Prep (Unit 3) Aim High (A-Level)Yasser MohamedNo ratings yet
- 5 Integumentary SystemDocument67 pages5 Integumentary SystemchelsealivesforeverNo ratings yet
- English DWDM 2022 Book 7 Int Sample PagesDocument7 pagesEnglish DWDM 2022 Book 7 Int Sample PagesLuiz Roberto MartinesNo ratings yet
- CaffeineDocument2 pagesCaffeineSaini Malkeet100% (1)
- Ceramic Foam Filter PDFDocument4 pagesCeramic Foam Filter PDFPaen ZulkifliNo ratings yet
- Modern tobacco marketing fundamentalsDocument6 pagesModern tobacco marketing fundamentalsAbdus SalamNo ratings yet
- Child Rights and Juvenile Justice in India MedhaDocument23 pagesChild Rights and Juvenile Justice in India MedhaLaw ColloquyNo ratings yet
- Hatch-Slack PathwayDocument10 pagesHatch-Slack Pathwaychurail khanNo ratings yet
- Mall Tenant Design ManualDocument52 pagesMall Tenant Design ManualWaleed AliNo ratings yet
- Jane Austen's Literary StyleDocument4 pagesJane Austen's Literary StyleGulfam RashidNo ratings yet
- CAE Proposal Expressions and SamplesDocument6 pagesCAE Proposal Expressions and SamplesKimLiênNguyễnNo ratings yet
- Paths To Mastery Lesson 17 Speaking Tips From Dale Carnegie The Quick and Easy Way To Effective SpeakingDocument10 pagesPaths To Mastery Lesson 17 Speaking Tips From Dale Carnegie The Quick and Easy Way To Effective SpeakingZelic NguyenNo ratings yet
- Route StructureDocument3 pagesRoute StructureAndrei Gideon ReyesNo ratings yet
- Syllabus Changes: March 2021 Version 1Document20 pagesSyllabus Changes: March 2021 Version 1Azul IrlaundeNo ratings yet
- KCIS 2nd Semester Science Fair ProjectsDocument2 pagesKCIS 2nd Semester Science Fair ProjectsCaT BlAcKNo ratings yet
- Nilai Murni PKN XII Mipa 3Document8 pagesNilai Murni PKN XII Mipa 3ilmi hamdinNo ratings yet
- Guide to Preparing STEM Fellowship ApplicationsDocument14 pagesGuide to Preparing STEM Fellowship ApplicationsNurrahmiNo ratings yet
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineFrom EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineRating: 4 out of 5 stars4/5 (17)
- Superlative: The Biology of ExtremesFrom EverandSuperlative: The Biology of ExtremesRating: 4.5 out of 5 stars4.5/5 (51)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Eels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishFrom EverandEels: An Exploration, from New Zealand to the Sargasso, of the World's Most Mysterious FishRating: 4 out of 5 stars4/5 (30)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Human Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesFrom EverandHuman Errors: A Panorama of Our Glitches, from Pointless Bones to Broken GenesRating: 3.5 out of 5 stars3.5/5 (56)
- Darwin's Dangerous Idea: Evolution and the Meaning of LifeFrom EverandDarwin's Dangerous Idea: Evolution and the Meaning of LifeRating: 4 out of 5 stars4/5 (523)
- The Mind & The Brain: Neuroplasticity and the Power of Mental ForceFrom EverandThe Mind & The Brain: Neuroplasticity and the Power of Mental ForceNo ratings yet
- The Dragons of Eden: Speculations on the Evolution of Human IntelligenceFrom EverandThe Dragons of Eden: Speculations on the Evolution of Human IntelligenceRating: 4 out of 5 stars4/5 (632)