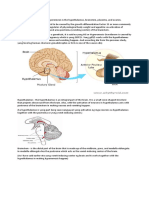
You might also like
- What Is Attachment ParentingDocument24 pagesWhat Is Attachment Parentingrelax5810No ratings yet
- Gyn MCQs PRIMEsDocument40 pagesGyn MCQs PRIMEssk100% (2)
- Reproductive System QuizDocument3 pagesReproductive System QuizJONEL FONTILLASNo ratings yet
- NCP Imbalanced Nutrition IUGR PatientDocument2 pagesNCP Imbalanced Nutrition IUGR Patientanreilegarde100% (10)
- Abnormal LabourDocument15 pagesAbnormal LabourAshish GuptaNo ratings yet
- General and Comparative Endocrinology: M. Gupta, S.S. Dangi, G. Singh, M. SarkarDocument9 pagesGeneral and Comparative Endocrinology: M. Gupta, S.S. Dangi, G. Singh, M. SarkarTeodora OnofreiNo ratings yet
- Jcem 1476Document6 pagesJcem 1476Teodora OnofreiNo ratings yet
- Fendo 10 00777Document14 pagesFendo 10 00777Fatima 999No ratings yet
- Uterine Adenocarcinoma in Mice Treated NDocument5 pagesUterine Adenocarcinoma in Mice Treated NAmeer MohammadNo ratings yet
- An Overview On Functional Causes of Infertility in CowsDocument6 pagesAn Overview On Functional Causes of Infertility in CowsAh MagdyNo ratings yet
- (17417899 - Reproduction) Role of Ghrelin in Fertilization, Early Embryo Development, and Implantation PeriodsDocument9 pages(17417899 - Reproduction) Role of Ghrelin in Fertilization, Early Embryo Development, and Implantation PeriodsMarco AsprónNo ratings yet
- Int J Fertil Steril 12 37Document6 pagesInt J Fertil Steril 12 37FarizNo ratings yet
- Embryo Endometrial Interaction Associated With The Location of The Embryo During The Mobility Phase in MaresDocument13 pagesEmbryo Endometrial Interaction Associated With The Location of The Embryo During The Mobility Phase in Marespazprieto24No ratings yet
- Recent Advances in Nutritional Sciences: Maternal Nutrition and Fetal DevelopmentDocument4 pagesRecent Advances in Nutritional Sciences: Maternal Nutrition and Fetal Developmentomarmeftah838No ratings yet
- It Takes Two To TangoDocument3 pagesIt Takes Two To Tangoaftab khanNo ratings yet
- 4 - Glutaminas y La Respuesta A La URP - Proteina Del Estres (Ingles)Document7 pages4 - Glutaminas y La Respuesta A La URP - Proteina Del Estres (Ingles)luis mozoNo ratings yet
- Theriogenology: SciencedirectDocument6 pagesTheriogenology: SciencedirectGustav MolMedNo ratings yet
- 2022 Papers Merged 58 70Document13 pages2022 Papers Merged 58 70Yeannie YapNo ratings yet
- Adgent Et Al - 2018 - A Longitudinal Study of Estrogen-Responsive Tissues and Hormone ConcentrationsDocument11 pagesAdgent Et Al - 2018 - A Longitudinal Study of Estrogen-Responsive Tissues and Hormone ConcentrationsBerry BancinNo ratings yet
- Reproductive Toxicology: SciencedirectDocument8 pagesReproductive Toxicology: SciencedirectBianca MickaelaNo ratings yet
- Muller Et Al-2013 PDFDocument9 pagesMuller Et Al-2013 PDFantoniosippelNo ratings yet
- 1 s2.0 S0015028213003993 MainDocument10 pages1 s2.0 S0015028213003993 MainMarianaafiatiNo ratings yet
- Ajuogu Etal 2019 PDFDocument7 pagesAjuogu Etal 2019 PDFAjuogu PeterNo ratings yet
- 1479 6805 Joe 21 0449Document12 pages1479 6805 Joe 21 0449Salim MichaelNo ratings yet
- Ajpendo 00007 2019Document13 pagesAjpendo 00007 2019Ali MazidiNo ratings yet
- Soybean-Derived Phytoestrogens Regulate Prostaglandin Secretion in Endometrium During Cattle Estrous Cycle and Early PregnancyDocument11 pagesSoybean-Derived Phytoestrogens Regulate Prostaglandin Secretion in Endometrium During Cattle Estrous Cycle and Early Pregnancysirous1No ratings yet
- Phytoestrogens The Natural Selective EstDocument5 pagesPhytoestrogens The Natural Selective EstronniemonsterNo ratings yet
- 1 s2.0 S0093691X22004472 MainDocument9 pages1 s2.0 S0093691X22004472 MainYuli CruzNo ratings yet
- Content ServerDocument8 pagesContent ServerMarco AsprónNo ratings yet
- Effect Of Prostaglandin F2Α And Gnrh Administration At The Time Of Artificial Insemination On Reproductive Performance Of Dairy CowsDocument6 pagesEffect Of Prostaglandin F2Α And Gnrh Administration At The Time Of Artificial Insemination On Reproductive Performance Of Dairy Cowsalfriyan MelatiNo ratings yet
- (14796805 - Journal of Endocrinology) Resistin Expression and Plasma Concentration Peak at Different Times During Pregnancy in RatsDocument9 pages(14796805 - Journal of Endocrinology) Resistin Expression and Plasma Concentration Peak at Different Times During Pregnancy in RatsDendrita DospuntoceroNo ratings yet
- Main - 10 38 06Document14 pagesMain - 10 38 06Yavuz CarasoNo ratings yet
- Mechanisms Underlying Reduced Fertility in Anovular Dairy CowsDocument9 pagesMechanisms Underlying Reduced Fertility in Anovular Dairy CowsIoana CriveiNo ratings yet
- Domestic Animal EndocrinologyDocument9 pagesDomestic Animal EndocrinologyJose Luis Ponce CovarrubiasNo ratings yet
- Ijms 22 11059Document36 pagesIjms 22 11059Teodora OnofreiNo ratings yet
- Reproduction: Reproductive Consequences of Developmental Phytoestrogen ExposureDocument14 pagesReproduction: Reproductive Consequences of Developmental Phytoestrogen ExposureAdib MustofaNo ratings yet
- A:ACEP 疗法Dr.SherDocument6 pagesA:ACEP 疗法Dr.Shermeltwithsnow163.comNo ratings yet
- Physiological Functions of Ghrelin:: A B, C DDocument5 pagesPhysiological Functions of Ghrelin:: A B, C DHR BajwaNo ratings yet
- Lahore StudyDocument5 pagesLahore StudyFawad YousufNo ratings yet
- Investigation of Irisin's Role in Pubertal Onset Physiology in Female RatsDocument6 pagesInvestigation of Irisin's Role in Pubertal Onset Physiology in Female RatsonderhuseyinbasNo ratings yet
- Fakriadis2018 GnRHa MaturationDocument10 pagesFakriadis2018 GnRHa Maturationanon_509992240No ratings yet
- 2015 Embryos, DOHaD and David BarkerDocument7 pages2015 Embryos, DOHaD and David BarkerNguyễn Tiến HồngNo ratings yet
- Antonio GonzálezDocument8 pagesAntonio GonzálezJose Luis Ponce CovarrubiasNo ratings yet
- Reproduccion en CrustaceosDocument10 pagesReproduccion en CrustaceosPabloVejarNo ratings yet
- Varuzza Et Al-2019-Environmental ToxicologyDocument10 pagesVaruzza Et Al-2019-Environmental ToxicologyCharles santos da costaNo ratings yet
- 2008 - QUIGLEY Et Al - Effect of Variable Long-Term Maternal Feed Allowance On The Development of The Ovine Placenta and FetusDocument10 pages2008 - QUIGLEY Et Al - Effect of Variable Long-Term Maternal Feed Allowance On The Development of The Ovine Placenta and FetusSamanta MonteiroNo ratings yet
- Etiology of HypospadiaDocument10 pagesEtiology of HypospadiaJemy IkkiNo ratings yet
- JC 2016-3452Document8 pagesJC 2016-3452Zoel NikonianNo ratings yet
- Assisted Reproductive Technologies in The Reproductive Management of Small RuminantsDocument10 pagesAssisted Reproductive Technologies in The Reproductive Management of Small RuminantsAlexander Cañar RomeroNo ratings yet
- Senapati2018 Article SuperovulationAltersTheExpressDocument10 pagesSenapati2018 Article SuperovulationAltersTheExpressInneke NoerNo ratings yet
- Gohlke 2005Document5 pagesGohlke 2005yulia fatma nstNo ratings yet
- The Science Behind 25 Years of Ovarian Stimulation For IVFDocument38 pagesThe Science Behind 25 Years of Ovarian Stimulation For IVFPatrick BayuNo ratings yet
- 1 Factores AutocrinosDocument5 pages1 Factores AutocrinosDanny ContrerasNo ratings yet
- Growth Hormone Deficiency in PseudohypoparathyroidDocument12 pagesGrowth Hormone Deficiency in Pseudohypoparathyroidradu nicolaeNo ratings yet
- Zhong Et Al., 2021Document8 pagesZhong Et Al., 2021Firat MeiyasaNo ratings yet
- Hormone Action in The Mammary Gland - C Brisken and B O Malley 2010Document15 pagesHormone Action in The Mammary Gland - C Brisken and B O Malley 2010tostonasNo ratings yet
- Animal Reproduction ScienceDocument8 pagesAnimal Reproduction ScienceCeliny CarvalhoNo ratings yet
- Docchio 2019 Influence of Nutrition Body ConditiDocument8 pagesDocchio 2019 Influence of Nutrition Body ConditiAna SandovalNo ratings yet
- Nutrition and Reproduction 1996 RobinsonDocument10 pagesNutrition and Reproduction 1996 RobinsonCanuto MunozNo ratings yet
- Placenta: SciencedirectDocument8 pagesPlacenta: SciencedirectMayra PereiraNo ratings yet
- (13142143 - Folia Medica) Evaluation of Teratogenic Activity of Antiepileptic Drug Lamotrigine in Mouse FetusesDocument6 pages(13142143 - Folia Medica) Evaluation of Teratogenic Activity of Antiepileptic Drug Lamotrigine in Mouse FetusesrayaimNo ratings yet
- Hormonal Control of Pig ReproductionDocument3 pagesHormonal Control of Pig ReproductionCarol OrequesNo ratings yet
- 2020 - Porcino - Oviducto Capacitación Espermática PDFDocument19 pages2020 - Porcino - Oviducto Capacitación Espermática PDFMarco Antonio AspronNo ratings yet
- BiotechnologyDocument9 pagesBiotechnologyÄsish Kumär SwainNo ratings yet
- Hormones and Embryonic Development: Advances in the BiosciencesFrom EverandHormones and Embryonic Development: Advances in the BiosciencesG. RaspéNo ratings yet
- Biological Beginnings - Learning ExercisesDocument4 pagesBiological Beginnings - Learning ExercisesNnieh KhuuNo ratings yet
- Discussion About Labor and DeliveryDocument3 pagesDiscussion About Labor and DeliveryStudent NurseNo ratings yet
- Hallazgos Normales y Anormales de La ColposDocument12 pagesHallazgos Normales y Anormales de La ColposAura ZuñigaNo ratings yet
- Pregnancy Induced HypertensionDocument43 pagesPregnancy Induced HypertensionDesty Adjaahh LaahhNo ratings yet
- Ge8 Guide Questions #11 (Natural Law)Document2 pagesGe8 Guide Questions #11 (Natural Law)jalil kablizoNo ratings yet
- FertilizationDocument63 pagesFertilizationDheressaaNo ratings yet
- PlatinaDocument61 pagesPlatinabhaveshsakariya555No ratings yet
- Torch Infection and Its Management With Ayurveda, An Indigenous MedicineDocument2 pagesTorch Infection and Its Management With Ayurveda, An Indigenous MedicineDhiraj BaksheNo ratings yet
- Gyne Prelims Finals ReviewerDocument27 pagesGyne Prelims Finals ReviewerLM N/ANo ratings yet
- Feminist CareDocument3 pagesFeminist Carelily bloomNo ratings yet
- Hanna Owo BlowjobDocument1 pageHanna Owo BlowjobVictorNo ratings yet
- Reproduction Physiology NotesDocument29 pagesReproduction Physiology NotesbilalNo ratings yet
- Anaphy & Pathophy (Hyperemesis)Document6 pagesAnaphy & Pathophy (Hyperemesis)Irish EspinosaNo ratings yet
- HSG PresentationDocument18 pagesHSG Presentationashikin92No ratings yet
- AM IV-Reading Material 2Document21 pagesAM IV-Reading Material 2Deshna JainNo ratings yet
- Placenta ExaminationDocument2 pagesPlacenta ExaminationSumit YadavNo ratings yet
- Module 2 Lesson 1 Physical and Sexual SelfDocument6 pagesModule 2 Lesson 1 Physical and Sexual SelfRyan Kenneth Oliver E. MacahipayNo ratings yet
- Family Planning/contraception Methods: Key FactsDocument4 pagesFamily Planning/contraception Methods: Key FactsCheesy YoguNo ratings yet
- Abortion in Kiambu County.Document30 pagesAbortion in Kiambu County.Joshua Marissah NaftaliNo ratings yet
- Nursing-Interventions MCN For Infertility MenDocument2 pagesNursing-Interventions MCN For Infertility MenRio DanteNo ratings yet
- Oym Test Planner - Phase 02Document5 pagesOym Test Planner - Phase 02ManishNo ratings yet
- Recombinant Human Luteinizing Hormone To Trigger Ovulation Randomized Controlled Dose Finding Pilot Study in Ovulation InductionDocument13 pagesRecombinant Human Luteinizing Hormone To Trigger Ovulation Randomized Controlled Dose Finding Pilot Study in Ovulation InductionDevansh ShethNo ratings yet
- In VitroDocument10 pagesIn VitroJulian David Ortiz LondoñoNo ratings yet
- Essay LIADocument7 pagesEssay LIAFerdi AnsyahNo ratings yet
- TOG Evolution in Screening For Down SyndromeDocument7 pagesTOG Evolution in Screening For Down SyndromeMarNo ratings yet