You might also like
- Structural and Functional Basis of Sars-Cov-2 Entry by Using Human Ace2Document21 pagesStructural and Functional Basis of Sars-Cov-2 Entry by Using Human Ace2rezaNo ratings yet
- Structure-Based Drug Discovery To Identify SARS-CoV2 Spike Protein ACE2 Interaction InhibitorsDocument20 pagesStructure-Based Drug Discovery To Identify SARS-CoV2 Spike Protein ACE2 Interaction Inhibitorsdenemebakalim72No ratings yet
- Microorganisms 10 00280 With CoverDocument8 pagesMicroorganisms 10 00280 With CoverEditions La LumièreNo ratings yet
- Sars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneDocument9 pagesSars-Cov-2 Receptor and Renin-Angiotensin System Regulation: Impact of Genetics Variants in Ace2 GeneInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Nac Papel Antioxidantes Cov-19 0221 FrattaDocument23 pagesNac Papel Antioxidantes Cov-19 0221 FrattaLuis C Ribon VNo ratings yet
- Corona ProfilDocument19 pagesCorona ProfilVincentius Michael WilliantoNo ratings yet
- Structural Basis of Receptor Recognition by Sars-Cov-2: ArticleDocument18 pagesStructural Basis of Receptor Recognition by Sars-Cov-2: Articlefelix sotoNo ratings yet
- Full PDFDocument8 pagesFull PDFYad Mohammad NazaryNo ratings yet
- Vaccine - RBD TArgetDocument9 pagesVaccine - RBD TArgetindahikasNo ratings yet
- ACE-2 em Cães e GatosDocument10 pagesACE-2 em Cães e GatosExpedito MaiaNo ratings yet
- 2021 Article 2395Document15 pages2021 Article 2395danyescalerapumas24No ratings yet
- Role of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19: Review Open AccessDocument10 pagesRole of Angiotensin-Converting Enzyme 2 (ACE2) in COVID-19: Review Open AccessjakpowerNo ratings yet
- Sars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinDocument10 pagesSars-Cov-2: Phylogenetic Status, Mutations and Therapeutic Research Based On Spike ProteinArley GutarraNo ratings yet
- 5400-Article Text-31589-2-10-20210820Document12 pages5400-Article Text-31589-2-10-20210820Voltaire Méndez RodríguezNo ratings yet
- Drug Sars-CovDocument20 pagesDrug Sars-CovSilvi Zakiyatul IlmiyahNo ratings yet
- Nuevo Coronavirus en MurcielagosDocument22 pagesNuevo Coronavirus en MurcielagosCarlos Vinicio Chiluisa GuachoNo ratings yet
- 1 s2.0 S2405844022021466 MainDocument8 pages1 s2.0 S2405844022021466 MainhogirimmunologyNo ratings yet
- nCoV - Med - 2020-06-05 - Enhanced Receptor Binding of SARS-CoV-2 Through Networks of Hydrogen-Bonding and Hydrophobic InteractionsDocument8 pagesnCoV - Med - 2020-06-05 - Enhanced Receptor Binding of SARS-CoV-2 Through Networks of Hydrogen-Bonding and Hydrophobic InteractionsKaroline MarxNo ratings yet
- Journal Pre-Proof: Infection, Genetics and EvolutionDocument29 pagesJournal Pre-Proof: Infection, Genetics and EvolutionMohammed Shuaib AhmedNo ratings yet
- Asesoria Articulo CientificoDocument8 pagesAsesoria Articulo CientificoWilson Ramos OlanoNo ratings yet
- A Novel Bat Coronavirus Closely Related To SARS-CoDocument14 pagesA Novel Bat Coronavirus Closely Related To SARS-CoIntzar MehdiNo ratings yet
- Structure-Altering Mutations of The Sars-Cov-2 Frameshifting Rna ElementDocument14 pagesStructure-Altering Mutations of The Sars-Cov-2 Frameshifting Rna ElementMd. Rafid HasanNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateainindyaNo ratings yet
- Journal Pre-Proof: Trends in ImmunologyDocument34 pagesJournal Pre-Proof: Trends in ImmunologyIntan AyuNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateZâna ZorilorNo ratings yet
- The Genetic Structure of Sars-Cov-2 Does Not Rule Out A Laboratory OriginDocument9 pagesThe Genetic Structure of Sars-Cov-2 Does Not Rule Out A Laboratory OriginNicholas SchmittNo ratings yet
- Journal Pre Proof Severe Respiratory SARDocument16 pagesJournal Pre Proof Severe Respiratory SARgygyNo ratings yet
- 1 s2.0 S0188440920302927 MainDocument5 pages1 s2.0 S0188440920302927 MainRheum RaponticumNo ratings yet
- A Review On Sars Cov 2 Virology, Pathophysiology, Animal Models, and Anti Viral InterventionsDocument22 pagesA Review On Sars Cov 2 Virology, Pathophysiology, Animal Models, and Anti Viral InterventionsalodiarkNo ratings yet
- Protocolo - 2022Document10 pagesProtocolo - 2022Carlos Alberto Salazar DuqueNo ratings yet
- Dholariya2021 Article NotableAndEmergingVariantsOfSADocument8 pagesDholariya2021 Article NotableAndEmergingVariantsOfSAA.R MaheshNo ratings yet
- Covid-19 Science Report:: Pathogenesis and Host Immune Response To Sars-Cov-2Document12 pagesCovid-19 Science Report:: Pathogenesis and Host Immune Response To Sars-Cov-2ChavdarNo ratings yet
- COVID-19: Pathophysiology, Diagnosis, Complications and Investigational TherapeuticsDocument8 pagesCOVID-19: Pathophysiology, Diagnosis, Complications and Investigational TherapeuticsHeidiNo ratings yet
- How Does Sars-Cov-2 Cause Covid-19?: The Viral Receptor On Human Cells Plays A Critical Role in Disease ProgressionDocument3 pagesHow Does Sars-Cov-2 Cause Covid-19?: The Viral Receptor On Human Cells Plays A Critical Role in Disease ProgressionMiguel Chimal MuñozNo ratings yet
- Crystal Structure of SARS CoV 2 Nucleocapsid Protein RNA - 2020 - Acta PharmaceDocument11 pagesCrystal Structure of SARS CoV 2 Nucleocapsid Protein RNA - 2020 - Acta PharmaceMohammed Shuaib AhmedNo ratings yet
- Sars Cov2 Antigen Presentation Process by Immune Cells: Immunology Assignment - 2Document17 pagesSars Cov2 Antigen Presentation Process by Immune Cells: Immunology Assignment - 2azeema fatimaNo ratings yet
- European Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilDocument11 pagesEuropean Journal of Pharmacology: Gayathri Krishna, Vinod Soman Pillai, Mohanan Valiya VeettilautomationenggNo ratings yet
- Reviews: Mechanisms of Sars-Cov-2 Entry Into CellsDocument18 pagesReviews: Mechanisms of Sars-Cov-2 Entry Into CellsRTEFG DFGJUNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateArifin SiregarNo ratings yet
- Sengupta & Dutta, 2022 - NAC & COVID-1Document6 pagesSengupta & Dutta, 2022 - NAC & COVID-1Dr.Pallav SenguptaNo ratings yet
- Gene of The Month: The 2019-Ncov/Sars - Cov-2 Novel Coronavirus Spike ProteinDocument4 pagesGene of The Month: The 2019-Ncov/Sars - Cov-2 Novel Coronavirus Spike ProteinNandagopal PaneerselvamNo ratings yet
- Journal of Infection and Public HealthDocument10 pagesJournal of Infection and Public Healthtuanda222No ratings yet
- Molecular Mechanism of Evolution and Human Infection With The Novel Coronavirus (2019-Ncov)Document19 pagesMolecular Mechanism of Evolution and Human Infection With The Novel Coronavirus (2019-Ncov)Eneko AlfandegaNo ratings yet
- The Proximal Origin of Sars-Cov-2Document16 pagesThe Proximal Origin of Sars-Cov-2perete69No ratings yet
- Natrulazing AntibodiDocument6 pagesNatrulazing Antibodidita novia maharaniNo ratings yet
- Journal Pre-Proof: European Journal of Internal MedicineDocument30 pagesJournal Pre-Proof: European Journal of Internal MedicineNabil Sangga BuanaNo ratings yet
- Jurnal Sinovac 2Document6 pagesJurnal Sinovac 2Delapan SembilanNo ratings yet
- Zerospike Project BrochureDocument24 pagesZerospike Project Brochurestephen.cordnerNo ratings yet
- Virology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateDocument6 pagesVirology, Transmission, and Pathogenesis of Sars-Cov-2: Clinical UpdateRocio GmarNo ratings yet
- Brojakowska 2020Document11 pagesBrojakowska 2020Anett Pappné LeppNo ratings yet
- Detection COVID19Document18 pagesDetection COVID19Luis KrNo ratings yet
- Kelor DocingDocument12 pagesKelor DocingtridewantiwNo ratings yet
- Journal of Structural Biology: X: A B C C D CDocument7 pagesJournal of Structural Biology: X: A B C C D CDaniel E. Zarcos LópezNo ratings yet
- Journal Pre-Proof: Clinical Microbiology and InfectionDocument23 pagesJournal Pre-Proof: Clinical Microbiology and Infectionsameer sahaanNo ratings yet
- Diarrhea During COVID-19 InfectionDocument11 pagesDiarrhea During COVID-19 InfectionMan'SzAr'diAnSyAhNo ratings yet
- Neutralizing Antibodies Against SARS-CoV-2 CurrentDocument15 pagesNeutralizing Antibodies Against SARS-CoV-2 CurrentAntonius SimangunsongNo ratings yet
- Stawiski 2020 Human - ACE2 - Receptor - Polymorphisms - Predict - SARS-CoVDocument33 pagesStawiski 2020 Human - ACE2 - Receptor - Polymorphisms - Predict - SARS-CoVGolden Era BookwormNo ratings yet
- El Origen Del Covid19Document5 pagesEl Origen Del Covid19Herrera NataliaNo ratings yet
- Epid Pubmed 1 (Bab2) .En - IdDocument11 pagesEpid Pubmed 1 (Bab2) .En - IdSHAFIYA ADELINANo ratings yet
- History of Cell: Robert HookeDocument4 pagesHistory of Cell: Robert HookeJenny HermosadoNo ratings yet
- Regents Biology Homework Packet Unit 5: Energy in A Cell Photosynthesis & Cellular RespirationDocument15 pagesRegents Biology Homework Packet Unit 5: Energy in A Cell Photosynthesis & Cellular RespirationHakan Alkan0% (1)
- District Resource Centre Mahbubnagar: Section - A (Short Essay Type Questions)Document2 pagesDistrict Resource Centre Mahbubnagar: Section - A (Short Essay Type Questions)tadepalli patanjaliNo ratings yet
- Molecular Biology Previous Year QuestionsDocument7 pagesMolecular Biology Previous Year QuestionsKrutika SNo ratings yet
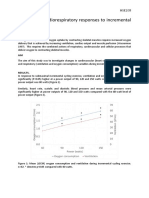
- Practical 4: Cardiorespiratory Responses To Incremental ExerciseDocument4 pagesPractical 4: Cardiorespiratory Responses To Incremental ExercisePATRICK OTIATONo ratings yet
- Vincento Frazier - Gizmos Lab - Protein SynthesisDocument6 pagesVincento Frazier - Gizmos Lab - Protein SynthesisToxic KikiNo ratings yet
- ChromoTek RFP BoosterDocument1 pageChromoTek RFP BoosterAlleleBiotechNo ratings yet
- Life Science KsetDocument16 pagesLife Science KsetRaghunathNo ratings yet
- IB DP Biology 1.3 Membrane Structure QuestionDocument15 pagesIB DP Biology 1.3 Membrane Structure QuestionikaNo ratings yet
- Viruses 08 00210Document13 pagesViruses 08 00210Hany ZutanNo ratings yet
- SB2a Mitosis 1. SB2b Growth in Animals SB2c Growth in PlantsDocument4 pagesSB2a Mitosis 1. SB2b Growth in Animals SB2c Growth in PlantsSiaNo ratings yet
- ACROBiosystems Product CatalogueDocument40 pagesACROBiosystems Product CatalogueArunee HansawongsakulNo ratings yet
- Fluorescence Microscopy FinalDocument45 pagesFluorescence Microscopy Finalapi-3845444100% (1)
- Cell Organelle Structure and Function Dhruv Tejusinghani P1Document6 pagesCell Organelle Structure and Function Dhruv Tejusinghani P1GamingWithDhruv100% (2)
- Sino Biological Inc. 2013 Product CatalogDocument181 pagesSino Biological Inc. 2013 Product CatalogMegan MckinneyNo ratings yet
- Heredit Y: Inheritance and Variation of TraitsDocument42 pagesHeredit Y: Inheritance and Variation of TraitsMae RicañaNo ratings yet
- Lipid AssignmentDocument4 pagesLipid AssignmentZainab AliNo ratings yet
- Botany ReviewerDocument10 pagesBotany ReviewerJARISSA DIZON GALLAZANo ratings yet
- AS Biology Unit1 Notes by Stafford Valentine ReddenDocument49 pagesAS Biology Unit1 Notes by Stafford Valentine ReddenAyaz Nasih88% (8)
- Biochemistry of The Blood Coagulation SystemDocument4 pagesBiochemistry of The Blood Coagulation Systemabc007200100% (1)
- pFUSE PDFDocument6 pagespFUSE PDFlaurakaiohNo ratings yet
- Histology & Its Methods of StudyDocument5 pagesHistology & Its Methods of StudyByron MacalintalNo ratings yet
- AS-Level Biology (New Spec) NotesDocument162 pagesAS-Level Biology (New Spec) NotesBlessing100% (1)
- Unit 6 & 7 Cell EnergyDocument17 pagesUnit 6 & 7 Cell EnergyRut ChristineNo ratings yet
- 5 PP Metabolism EnzymesDocument40 pages5 PP Metabolism EnzymesJharaNo ratings yet
- UWorld Biochem ReviewDocument1 pageUWorld Biochem ReviewvsalardaNo ratings yet
- @MedicalBooksStore 2014 ContraceptionDocument315 pages@MedicalBooksStore 2014 ContraceptionBernardo Fernandez SalazarNo ratings yet
- Lab 1 AnatomyDocument5 pagesLab 1 AnatomyNimra Shahnaz Khadim HussainNo ratings yet
- Copyright © The McGraw-Hill Companies, Inc. Permission Required For ReproductionDocument33 pagesCopyright © The McGraw-Hill Companies, Inc. Permission Required For ReproductionKvaleramNo ratings yet
- Handbook On Metalloproteins Berti (Dekker 2001) PDFDocument1,210 pagesHandbook On Metalloproteins Berti (Dekker 2001) PDFAnamarijaNo ratings yet
- Grokking Algorithms: An illustrated guide for programmers and other curious peopleFrom EverandGrokking Algorithms: An illustrated guide for programmers and other curious peopleRating: 4 out of 5 stars4/5 (16)
- Laws of UX: Using Psychology to Design Better Products & ServicesFrom EverandLaws of UX: Using Psychology to Design Better Products & ServicesRating: 5 out of 5 stars5/5 (9)
- The Internet Con: How to Seize the Means of ComputationFrom EverandThe Internet Con: How to Seize the Means of ComputationRating: 5 out of 5 stars5/5 (6)
- The Designer’s Guide to Figma: Master Prototyping, Collaboration, Handoff, and WorkflowFrom EverandThe Designer’s Guide to Figma: Master Prototyping, Collaboration, Handoff, and WorkflowNo ratings yet
- Branding: What You Need to Know About Building a Personal Brand and Growing Your Small Business Using Social Media Marketing and Offline Guerrilla TacticsFrom EverandBranding: What You Need to Know About Building a Personal Brand and Growing Your Small Business Using Social Media Marketing and Offline Guerrilla TacticsRating: 5 out of 5 stars5/5 (32)
- Social Media Marketing 2024, 2025: Build Your Business, Skyrocket in Passive Income, Stop Working a 9-5 Lifestyle, True Online Working from HomeFrom EverandSocial Media Marketing 2024, 2025: Build Your Business, Skyrocket in Passive Income, Stop Working a 9-5 Lifestyle, True Online Working from HomeNo ratings yet
- Introduction to Internet Scams and Fraud: Credit Card Theft, Work-At-Home Scams and Lottery ScamsFrom EverandIntroduction to Internet Scams and Fraud: Credit Card Theft, Work-At-Home Scams and Lottery ScamsRating: 4 out of 5 stars4/5 (6)
- Tor Darknet Bundle (5 in 1): Master the Art of InvisibilityFrom EverandTor Darknet Bundle (5 in 1): Master the Art of InvisibilityRating: 4.5 out of 5 stars4.5/5 (5)
- Practical Industrial Cybersecurity: ICS, Industry 4.0, and IIoTFrom EverandPractical Industrial Cybersecurity: ICS, Industry 4.0, and IIoTNo ratings yet
- Ten Arguments for Deleting Your Social Media Accounts Right NowFrom EverandTen Arguments for Deleting Your Social Media Accounts Right NowRating: 4 out of 5 stars4/5 (388)
- Facing Cyber Threats Head On: Protecting Yourself and Your BusinessFrom EverandFacing Cyber Threats Head On: Protecting Yourself and Your BusinessRating: 4.5 out of 5 stars4.5/5 (27)
- Evaluation of Some SMS Verification Services and Virtual Credit Cards Services for Online Accounts VerificationsFrom EverandEvaluation of Some SMS Verification Services and Virtual Credit Cards Services for Online Accounts VerificationsRating: 5 out of 5 stars5/5 (1)
- More Porn - Faster!: 50 Tips & Tools for Faster and More Efficient Porn BrowsingFrom EverandMore Porn - Faster!: 50 Tips & Tools for Faster and More Efficient Porn BrowsingRating: 3.5 out of 5 stars3.5/5 (24)
- The Digital Marketing Handbook: A Step-By-Step Guide to Creating Websites That SellFrom EverandThe Digital Marketing Handbook: A Step-By-Step Guide to Creating Websites That SellRating: 5 out of 5 stars5/5 (6)
- Python for Beginners: The 1 Day Crash Course For Python Programming In The Real WorldFrom EverandPython for Beginners: The 1 Day Crash Course For Python Programming In The Real WorldNo ratings yet
- The Ultimate LinkedIn Sales Guide: How to Use Digital and Social Selling to Turn LinkedIn into a Lead, Sales and Revenue Generating MachineFrom EverandThe Ultimate LinkedIn Sales Guide: How to Use Digital and Social Selling to Turn LinkedIn into a Lead, Sales and Revenue Generating MachineNo ratings yet
- The Dark Net: Inside the Digital UnderworldFrom EverandThe Dark Net: Inside the Digital UnderworldRating: 3.5 out of 5 stars3.5/5 (104)
- React.js Design Patterns: Learn how to build scalable React apps with ease (English Edition)From EverandReact.js Design Patterns: Learn how to build scalable React apps with ease (English Edition)No ratings yet
- Evaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsFrom EverandEvaluation of Some Websites that Offer Virtual Phone Numbers for SMS Reception and Websites to Obtain Virtual Debit/Credit Cards for Online Accounts VerificationsRating: 5 out of 5 stars5/5 (1)
- The YouTube Formula: How Anyone Can Unlock the Algorithm to Drive Views, Build an Audience, and Grow RevenueFrom EverandThe YouTube Formula: How Anyone Can Unlock the Algorithm to Drive Views, Build an Audience, and Grow RevenueRating: 5 out of 5 stars5/5 (33)