You might also like
- The Sympathetic Nervous System: The Muse of Primary HypertensionDocument6 pagesThe Sympathetic Nervous System: The Muse of Primary HypertensionRaihan NahrizaNo ratings yet
- Adrenergic Response To Morphine Diazepam.7Document6 pagesAdrenergic Response To Morphine Diazepam.7juan david castillo campoNo ratings yet
- Pulse Rate Variability Is Not A Surrogate For Heart Rate VariabilityDocument7 pagesPulse Rate Variability Is Not A Surrogate For Heart Rate VariabilityFiras ZakNo ratings yet
- L Malmqvist (ESPAÑOL PUMED)Document5 pagesL Malmqvist (ESPAÑOL PUMED)adr3an.tabaresNo ratings yet
- Time and Frequency Domain Analysis of Heart Rate VariabilityDocument7 pagesTime and Frequency Domain Analysis of Heart Rate VariabilityArchana TatavartiNo ratings yet
- MilicevicDocument6 pagesMilicevicKlaudijus VitkauskasNo ratings yet
- Net Notes CardioDocument9 pagesNet Notes CardioJess PeltraNo ratings yet
- The Central Renin-Angiotensin System and Sympathetic Nerve Activity in Chronic Heart FailureDocument13 pagesThe Central Renin-Angiotensin System and Sympathetic Nerve Activity in Chronic Heart FailureHefie RahmaniarNo ratings yet
- Effects of Saffan Cardiopulmonary in Healthy: FunctionDocument4 pagesEffects of Saffan Cardiopulmonary in Healthy: FunctionHocicos Y PatasNo ratings yet
- Montano 1992Document11 pagesMontano 1992Mary Marlene Tarazona MolinaNo ratings yet
- 1 s2.0 S073510970800778X MainDocument2 pages1 s2.0 S073510970800778X MainvanathyNo ratings yet
- Paper Inmunosensores para Tratamientos Del CancerDocument7 pagesPaper Inmunosensores para Tratamientos Del CancerMauro MarchettoNo ratings yet
- Heart Rate Variability in Anaesthesiology - Narrative ReviewDocument8 pagesHeart Rate Variability in Anaesthesiology - Narrative ReviewvienvienkkNo ratings yet
- Heart Rate Variability: A Diagnostic and Prognostic Tool in Anesthesia and Intensive CareDocument15 pagesHeart Rate Variability: A Diagnostic and Prognostic Tool in Anesthesia and Intensive CarenrlaprilianiNo ratings yet
- Cardiac Sympathovagal Balance During Sleep Apnea Episodes: Vanninen, A. Tuunainen, M. Kansanen,? Uusitupas and LansimiesDocument8 pagesCardiac Sympathovagal Balance During Sleep Apnea Episodes: Vanninen, A. Tuunainen, M. Kansanen,? Uusitupas and LansimiesaudiNo ratings yet
- Overview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive TherapyDocument11 pagesOverview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive Therapyحمزة جمالNo ratings yet
- Overview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive TherapyDocument11 pagesOverview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive Therapyحمزة جمالNo ratings yet
- Overview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive TherapyDocument11 pagesOverview of Al-Adrenoceptor Antagonism and Recent Advances in Hypertensive Therapyحمزة جمالNo ratings yet
- Ajpheart 1985 249 4 h867Document9 pagesAjpheart 1985 249 4 h867Rakibul HasanNo ratings yet
- Sympathetic Modulation by Antihypertensive DrugsDocument3 pagesSympathetic Modulation by Antihypertensive Drugsthomas albertNo ratings yet
- 445Document8 pages445teodoraNo ratings yet
- Heart Rate Variability Measurement and CDocument14 pagesHeart Rate Variability Measurement and CMary Marlene Tarazona MolinaNo ratings yet
- Clinical Cardiology - August 1990 - Malik - Heart Rate VariabilityDocument7 pagesClinical Cardiology - August 1990 - Malik - Heart Rate VariabilityP AlexiaNo ratings yet
- Pi Is 0007091217435811Document6 pagesPi Is 0007091217435811Julio AltamiranoNo ratings yet
- Effects of Hypertonic Saline On Myocardial Contractility in AnaesthetizedDocument7 pagesEffects of Hypertonic Saline On Myocardial Contractility in AnaesthetizedliyaNo ratings yet
- Passive Leg Raising and Fluid Responsiveness Monnet CCM 2006Document6 pagesPassive Leg Raising and Fluid Responsiveness Monnet CCM 2006Leiniker Navarro ReyNo ratings yet
- Noninvasive Autonomic and Hemodynamic Monitoring in Severe SepsisDocument9 pagesNoninvasive Autonomic and Hemodynamic Monitoring in Severe SepsisKamal NasirNo ratings yet
- Assessing Autonomic Function From Electrodermal Activity and Heart Rate Variability During Cold-Pressor Test and Emotional ChallengeDocument13 pagesAssessing Autonomic Function From Electrodermal Activity and Heart Rate Variability During Cold-Pressor Test and Emotional ChallengeMuh SutraNo ratings yet
- NT BNPDocument9 pagesNT BNPSagar MhetreNo ratings yet
- Clinical Implications of Intrarenal Hemodynamic Evaluation by Doppler Ultrasonography in Heart FailureDocument9 pagesClinical Implications of Intrarenal Hemodynamic Evaluation by Doppler Ultrasonography in Heart FailureRaul FernandoNo ratings yet
- Evaluation of The Relationship Between Peripheral Pulse Palpation and Doppler Systolic Blood Pressure in Dogs Presenting To An Emergency ServiceDocument6 pagesEvaluation of The Relationship Between Peripheral Pulse Palpation and Doppler Systolic Blood Pressure in Dogs Presenting To An Emergency ServiceYissette Bautista CarrilloNo ratings yet
- MainDocument11 pagesMainDhonz R AdiwaramanNo ratings yet
- Differences in Left Ventricular and Left Atrial FuDocument10 pagesDifferences in Left Ventricular and Left Atrial FueugeniaNo ratings yet
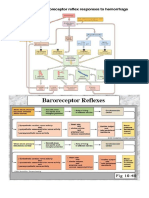
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionJoão Pedro Dantas AlkimimNo ratings yet
- Reiki1 Mackay NDocument6 pagesReiki1 Mackay NSoulyogaNo ratings yet
- Heart Rate Variability Evaluation in The Assessment of Cardiac Autonomic Neuropathy in Patients With Type 2 DiabetesDocument10 pagesHeart Rate Variability Evaluation in The Assessment of Cardiac Autonomic Neuropathy in Patients With Type 2 DiabetesPamelita Gutierrez PortelaNo ratings yet
- 11 - Arterial Blood Pressure RegulationDocument31 pages11 - Arterial Blood Pressure RegulationMumtaz MaulanaNo ratings yet
- Evaluation of Heart Rate Variability - UpToDateDocument31 pagesEvaluation of Heart Rate Variability - UpToDateGonçalo MirandaNo ratings yet
- Scale Exponents of Blood Pressure and Heart Rate During Autonomic Blockade As Assessed by Detrended Fluctuation AnalysisDocument15 pagesScale Exponents of Blood Pressure and Heart Rate During Autonomic Blockade As Assessed by Detrended Fluctuation Analysispokolio silvaNo ratings yet
- Heart Rate Variability PDFDocument7 pagesHeart Rate Variability PDFmik1989No ratings yet
- Reflexes That Control Cardiovascular FunctionDocument11 pagesReflexes That Control Cardiovascular FunctionRaam NaikNo ratings yet
- Chronic Interactions Between Carotid Baroreceptors and Chemoreceptors in Obesity HypertensionDocument14 pagesChronic Interactions Between Carotid Baroreceptors and Chemoreceptors in Obesity HypertensionCaesar Catalin CaratasuNo ratings yet
- HEARTRATEVARIABILITYDocument8 pagesHEARTRATEVARIABILITYdenta aeNo ratings yet
- Acute Effect of Kapalbhati Yoga On Cardiac AutonomDocument7 pagesAcute Effect of Kapalbhati Yoga On Cardiac AutonomAkarat SivaphongthongchaiNo ratings yet
- Effects of Respiratory Interval On Vagal Modulation of Heart RateDocument8 pagesEffects of Respiratory Interval On Vagal Modulation of Heart RateGuillermo Castro ContardoNo ratings yet
- Cardiovascular Pleiotropic Effects of NUPDocument18 pagesCardiovascular Pleiotropic Effects of NUPВладимир ДружининNo ratings yet
- Hipotermi 9Document4 pagesHipotermi 9wahyuartyningsihNo ratings yet
- Div Class Title Decreased Heart Rate Variability During Emotion Regulation in Subjects at Risk For Psychopathology DivDocument9 pagesDiv Class Title Decreased Heart Rate Variability During Emotion Regulation in Subjects at Risk For Psychopathology DivP AlexiaNo ratings yet
- Interdependent Feedback Regulation of Breathing by The Carotid Bodies and The Retrotrapezoid NucleusDocument14 pagesInterdependent Feedback Regulation of Breathing by The Carotid Bodies and The Retrotrapezoid NucleusLezard DomiNo ratings yet
- New Pharmacologic Therapies For AHFDocument9 pagesNew Pharmacologic Therapies For AHFDaniel HernandezNo ratings yet
- 2000 Adrenergic and Reflex Abnormalities in Obesity-Related HypertensionDocument5 pages2000 Adrenergic and Reflex Abnormalities in Obesity-Related HypertensionFerroBemNo ratings yet
- JTCM 1 51Document6 pagesJTCM 1 51soylahijadeunvampiroNo ratings yet
- Characterization of Complex Heart Rate Dynamics and Their Pharmacological Disorders by Non-Linear Prediction and Special Data TransformationsDocument7 pagesCharacterization of Complex Heart Rate Dynamics and Their Pharmacological Disorders by Non-Linear Prediction and Special Data TransformationsAnaAAAMNo ratings yet
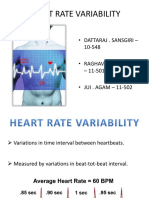
- Heart Rate Variability: - Dattaraj - Sansgiri - 10-548 - Raghav. Narasimhan - 11-501 - JUI - AGAM - 11-502Document27 pagesHeart Rate Variability: - Dattaraj - Sansgiri - 10-548 - Raghav. Narasimhan - 11-501 - JUI - AGAM - 11-502cloantafNo ratings yet
- CVS2Document10 pagesCVS2Ivana MilakovicNo ratings yet
- The Sympathetic Nervous System in Heart FailureDocument18 pagesThe Sympathetic Nervous System in Heart FailureCamilo VidalNo ratings yet
- PAM Metas Terapéuticas y Soporte FarmocológicoDocument9 pagesPAM Metas Terapéuticas y Soporte FarmocológicoHernando CastrillónNo ratings yet
- Howtotreatrightheart Failure - Tipsforclinicians IneverydaypracticeDocument11 pagesHowtotreatrightheart Failure - Tipsforclinicians IneverydaypracticeIsrael David Villalba FonsecaNo ratings yet
- Linear and Non-Linear Analyses of Heart Rate Variability: A MinireviewDocument9 pagesLinear and Non-Linear Analyses of Heart Rate Variability: A MinireviewMarkoCar16No ratings yet
- Geweke 1984Document10 pagesGeweke 1984Ivana MilakovicNo ratings yet
- Analysis of Capacity and Scalability of The LoRa Low Power Wide Area Network TechnologyDocument6 pagesAnalysis of Capacity and Scalability of The LoRa Low Power Wide Area Network TechnologyIvana MilakovicNo ratings yet
- CVS2Document10 pagesCVS2Ivana MilakovicNo ratings yet
- Cellular LTE-A Technologies For The Future Internet-of-Things: Physical Layer Features and ChallengesDocument28 pagesCellular LTE-A Technologies For The Future Internet-of-Things: Physical Layer Features and ChallengeshusseinelatarNo ratings yet
- JMater Chem BMagsensacceptedversionDocument41 pagesJMater Chem BMagsensacceptedversionIvana MilakovicNo ratings yet
- 46 Faes BiolCyb2013Document16 pages46 Faes BiolCyb2013Ivana MilakovicNo ratings yet
- Analysis of Capacity and Scalability of The LoRa Low Power Wide Area Network TechnologyDocument6 pagesAnalysis of Capacity and Scalability of The LoRa Low Power Wide Area Network TechnologyIvana MilakovicNo ratings yet
- Unit 3Document15 pagesUnit 3Neenu RajputNo ratings yet
- Lesson Plan-Pregnant WomenDocument5 pagesLesson Plan-Pregnant Womenapi-270972965No ratings yet
- Sattvika Dite-YakubDocument6 pagesSattvika Dite-Yakubyakubjacob100% (1)
- Individuals Trained To The Awareness Level Are Personnel Who Are Likely To Witness An Incident and Have Been Trained To Initiate Emergency Response byDocument21 pagesIndividuals Trained To The Awareness Level Are Personnel Who Are Likely To Witness An Incident and Have Been Trained To Initiate Emergency Response byacastillojose100% (1)
- 28 CryptorchidismDocument23 pages28 CryptorchidismFaisal Ibn MushtaqNo ratings yet
- CXC Csec English A June 2015 p1Document16 pagesCXC Csec English A June 2015 p1Rihanna Johnson100% (1)
- Tienchi Ginseng Panax Notoginseng San Qircljo PDFDocument4 pagesTienchi Ginseng Panax Notoginseng San Qircljo PDFTobiasenHolgersen89No ratings yet
- Postural DrainageDocument7 pagesPostural DrainagemohtishimNo ratings yet
- 48069B - Ferrofos 5260 (GB-ENG)Document6 pages48069B - Ferrofos 5260 (GB-ENG)danielNo ratings yet
- Surat Permintaan Obat Dinkes BaruDocument21 pagesSurat Permintaan Obat Dinkes BaruHafizh JhunkoNo ratings yet
- Test Bank For Mosbys Essentials For Nursing Assistants 4th Edition SorrentinoDocument24 pagesTest Bank For Mosbys Essentials For Nursing Assistants 4th Edition Sorrentinozacharymcleanpdqogjyfeb100% (47)
- Alice in Michigan: A Financial Hardship StudyDocument58 pagesAlice in Michigan: A Financial Hardship StudydaneNo ratings yet
- Spring Term Core A Formative AssessmentDocument9 pagesSpring Term Core A Formative Assessmentdiscord.tazzNo ratings yet
- Eritro Papulo Skuamosa 2021Document47 pagesEritro Papulo Skuamosa 2021Yolanda Agnesia Purba0% (1)
- Comparing and Contrasting Lady Macbeth and Ophelia From HamletDocument5 pagesComparing and Contrasting Lady Macbeth and Ophelia From HamletYashika MalhotraNo ratings yet
- Are You Planning To Buy or Rent A Home Built Before 1978?Document60 pagesAre You Planning To Buy or Rent A Home Built Before 1978?Maria FloresNo ratings yet
- EMP Procedure in MalaysiaDocument24 pagesEMP Procedure in Malaysialamkinpark3373No ratings yet
- Hizon Interview Group 2Document3 pagesHizon Interview Group 2Dan HizonNo ratings yet
- I-Stat 1 Quick Reference Guide 716942-00l CompressedDocument7 pagesI-Stat 1 Quick Reference Guide 716942-00l Compressedنجلاء الطلحيNo ratings yet
- Direct Black 22Document4 pagesDirect Black 22DHRUVNo ratings yet
- Approval Romania 2008Document27 pagesApproval Romania 2008Andrés Domínguez RuizNo ratings yet
- The Legal Bases of Special EducationDocument14 pagesThe Legal Bases of Special EducationMaria Dulcinea Basbas86% (7)
- Factories ActDocument64 pagesFactories ActKrishna Kaushik100% (1)
- Hyoscine ButylbromideDocument2 pagesHyoscine ButylbromideKenneth ColeNo ratings yet
- Eye Eye Assessment Assessment and Care and CareDocument126 pagesEye Eye Assessment Assessment and Care and CareBern NerquitNo ratings yet
- Contingency PlanDocument1 pageContingency PlanPramod Bodne100% (3)
- Common Skin Diseases in Africa: An Illustrated GuideDocument85 pagesCommon Skin Diseases in Africa: An Illustrated GuideJohn EnioladeNo ratings yet
- PMDC Community Medicine Mcqs PDFDocument280 pagesPMDC Community Medicine Mcqs PDFJay KNo ratings yet
- Drugs of Autonomic Nervous SystemDocument34 pagesDrugs of Autonomic Nervous SystemWyz Class100% (1)
- Whitesboro Isd - 1995 TEXAS SCHOOL SURVEY OF DRUG AND ALCOHOL USEDocument6 pagesWhitesboro Isd - 1995 TEXAS SCHOOL SURVEY OF DRUG AND ALCOHOL USETexas School Survey of Drug and Alcohol UseNo ratings yet