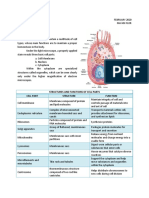
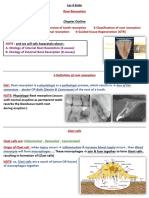
You might also like
- Pathway of The Pulp 10th EditionDocument1 pagePathway of The Pulp 10th Editionreal_septiady_madrid353225% (4)
- Gods WhispersDocument23 pagesGods WhispersRocíoEncarnaciónNo ratings yet
- GI Tract Histology SummaryDocument5 pagesGI Tract Histology SummaryFluffy_iceNo ratings yet
- 5.pulpotomy Medicaments Used in Deciduous Dentition An UpdateDocument18 pages5.pulpotomy Medicaments Used in Deciduous Dentition An UpdateRahma IskNo ratings yet
- Ceu Pre Board Exam 7 Ortho PedoDocument7 pagesCeu Pre Board Exam 7 Ortho PedoTara Lingating100% (2)
- Git Trans-Oral Cavity To StomachDocument6 pagesGit Trans-Oral Cavity To StomachHanako Sasaki AranillaNo ratings yet
- Experiment No. 10 Digestive System ObjectiveDocument5 pagesExperiment No. 10 Digestive System ObjectiveArman Renz PauloNo ratings yet
- Anaphy Notes Group 4Document21 pagesAnaphy Notes Group 4Purple BombNo ratings yet
- Medical Terminology and Anatomy For ICD-10 CodingDocument1 pageMedical Terminology and Anatomy For ICD-10 CodingsrdjanNo ratings yet
- Physiology Spring: Lecture 22 Study Guide: Chapter 16 The Digestive System 1 (P 565 - 597)Document12 pagesPhysiology Spring: Lecture 22 Study Guide: Chapter 16 The Digestive System 1 (P 565 - 597)Ngọc QuỳnhNo ratings yet
- Anaphylec Digestive System Finals GelaDocument20 pagesAnaphylec Digestive System Finals GelaPutri Oskieah MuhammadNo ratings yet
- Biology of Oral Mucosa and EsophagusDocument9 pagesBiology of Oral Mucosa and EsophagusJuan Sierra ZambranoNo ratings yet
- Histology - TissuesDocument8 pagesHistology - TissuesJm AshiiNo ratings yet
- 'Aliah's GIT & HB Systems1Document39 pages'Aliah's GIT & HB Systems1Luqman Al-Bashir FauziNo ratings yet
- Lab 4hzjDocument11 pagesLab 4hzj2cyvgpcs8zNo ratings yet
- Digestive System Part 1Document12 pagesDigestive System Part 1عبدالرحمن عدي عبدالفتاحNo ratings yet
- 10 Digestive System: C.T. Reyes, DVM, MSDocument26 pages10 Digestive System: C.T. Reyes, DVM, MSKent Solis VicenteNo ratings yet
- DIGESTIVEDocument21 pagesDIGESTIVEFuture TrekingNo ratings yet
- Practical Class 5Document73 pagesPractical Class 5Chahinez HabibNo ratings yet
- Digestive SystemDocument5 pagesDigestive SystemKate BambalanNo ratings yet
- And Descending Colon, Rectum, and Anal Canal. TheseDocument2 pagesAnd Descending Colon, Rectum, and Anal Canal. TheseDanielle Anne Zamora-Matillosa LambanNo ratings yet
- The Enteric Network - Interactions Between The Immune and Nervous Systems of The GutDocument17 pagesThe Enteric Network - Interactions Between The Immune and Nervous Systems of The GutMinjung KimNo ratings yet
- 6.1 Digestion and AbsorptionDocument18 pages6.1 Digestion and AbsorptionStacey Deo MakoyeNo ratings yet
- Tissue TransesDocument13 pagesTissue TransesBSMT1A ALMONIA, ERICKA ROSENo ratings yet
- Digestive System LecDocument13 pagesDigestive System LecMikaella ViadorNo ratings yet
- Lab Exercise No. 3Document3 pagesLab Exercise No. 3JOHN ISAAC BENITEZNo ratings yet
- Structural Organization in Animals Animal TissuesDocument3 pagesStructural Organization in Animals Animal TissuesHadiya FatimaNo ratings yet
- Activity 3Document6 pagesActivity 3Jefferson SociasNo ratings yet
- Stomach To Colon MircoDocument5 pagesStomach To Colon MircoLemuel ValerioNo ratings yet
- Anatomy IntroDocument4 pagesAnatomy IntroAlex HirschNo ratings yet
- Anatomy & Physiology Outlines - Finals (Chapter 16: Digestive)Document15 pagesAnatomy & Physiology Outlines - Finals (Chapter 16: Digestive)Dahrell Lopez BautistaNo ratings yet
- DLA 10 - Gastrointestinal Tract OverviewDocument13 pagesDLA 10 - Gastrointestinal Tract OverviewLloyd Bijuli-SinghNo ratings yet
- Digestive System: Abdominal QuadrantsDocument4 pagesDigestive System: Abdominal QuadrantsAngel Rose BrillanteNo ratings yet
- Histology of Oral Mucous Membrane and GingivaDocument86 pagesHistology of Oral Mucous Membrane and GingivahazeemmegahedNo ratings yet
- Digestive System: By: DhabaDocument84 pagesDigestive System: By: DhabanursefahussenNo ratings yet
- Tissues & Membranes: The Relationship Between Structure and Function of Epithelial TissuesDocument3 pagesTissues & Membranes: The Relationship Between Structure and Function of Epithelial Tissueskyla columnaNo ratings yet
- Mila Amor V. Reyes, MD, FPSP Anatomic and Clinical PathologistDocument41 pagesMila Amor V. Reyes, MD, FPSP Anatomic and Clinical PathologistFranco Rommel ReyesNo ratings yet
- W4 Tissue Level of OrgDocument13 pagesW4 Tissue Level of OrgWisNo ratings yet
- 9780198393511Document12 pages9780198393511Alexander CullenNo ratings yet
- Digestion and Absorption: Digestive ProcessDocument6 pagesDigestion and Absorption: Digestive ProcessJoao KasprowiczNo ratings yet
- Oral Mucous Membrane (Oral Mucosa)Document7 pagesOral Mucous Membrane (Oral Mucosa)روان امير غني عبد عونNo ratings yet
- Study Guide 23 - DIGESTIVE SYSTEMDocument33 pagesStudy Guide 23 - DIGESTIVE SYSTEMjed larsen capulong gavinoNo ratings yet
- Anaphy Digestive SystemDocument7 pagesAnaphy Digestive SystemShania ArevaloNo ratings yet
- Histology of Small Intestine 3Document17 pagesHistology of Small Intestine 3ShizraNo ratings yet
- Tissues and HistologyDocument8 pagesTissues and Histologycordero.kurt.bmlsNo ratings yet
- Maya Tejasari Histology Department Faculty of Medicine Bandung Islamic UniversityDocument65 pagesMaya Tejasari Histology Department Faculty of Medicine Bandung Islamic UniversitydeasyahNo ratings yet
- Histology AssignmentDocument6 pagesHistology AssignmentBryan LebasnonNo ratings yet
- Digestive SYSTEMDocument53 pagesDigestive SYSTEMPrincess AnnNo ratings yet
- Connective TissuesDocument6 pagesConnective TissuesPenelopeNo ratings yet
- Repaso Histologi ADocument132 pagesRepaso Histologi AMichael Rivera MoralesNo ratings yet
- Gen BioDocument10 pagesGen BioChristian RoqueNo ratings yet
- Jejunum (Small Intestine) Gastrointestinal TractDocument1 pageJejunum (Small Intestine) Gastrointestinal TractSabrina NodariNo ratings yet
- The Digestive System IDocument55 pagesThe Digestive System IYousef KhallafNo ratings yet
- Oral Mucosa: (Ten Cates)Document14 pagesOral Mucosa: (Ten Cates)Vanshika SethiNo ratings yet
- Module 5 - Digestive and Urinary SystemDocument31 pagesModule 5 - Digestive and Urinary SystemCaityNo ratings yet
- Chapter Twenty-Three The Digestive SystemDocument19 pagesChapter Twenty-Three The Digestive Systemjoni kashnerNo ratings yet
- TopicDocument3 pagesTopicShane VillaluzNo ratings yet
- Histology Notes 2Document13 pagesHistology Notes 2Lukas BüchlerNo ratings yet
- BIO122 - CHAPTER 7 Part 1Document53 pagesBIO122 - CHAPTER 7 Part 1lili100% (1)
- Esophagus Gastrointestinal TractDocument1 pageEsophagus Gastrointestinal TractpaolacsuarezaNo ratings yet
- The Small Intestine PDFDocument50 pagesThe Small Intestine PDFkhaledNo ratings yet
- Epithelial Tissues 2024-01Document45 pagesEpithelial Tissues 2024-01jcrosa137No ratings yet
- Treatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsFrom EverandTreatise on the Anatomy and Physiology of the Mucous Membranes: With Illustrative Pathological ObservationsNo ratings yet
- IndianJDentSci12268-3597318 095933Document5 pagesIndianJDentSci12268-3597318 095933TriptiLath DentalNo ratings yet
- An Overview of Oral TissuesDocument4 pagesAn Overview of Oral TissuesRenukaChauveNo ratings yet
- Pediatric DentistryDocument70 pagesPediatric Dentistrysruthy484100% (3)
- Sequelae of Dental CariesDocument20 pagesSequelae of Dental Cariesimi4100% (2)
- Pulp Therapy For Primary and Immature Permanent TeethDocument11 pagesPulp Therapy For Primary and Immature Permanent TeethPrabhjot KaurNo ratings yet
- Bleaching of Severely Darkened Nonvital Tooth Case - 48 Months Clinical ControlDocument8 pagesBleaching of Severely Darkened Nonvital Tooth Case - 48 Months Clinical ControlPriska talithaNo ratings yet
- Dental Traumatology-Medicolegal Aspects (Indian Perspective)Document6 pagesDental Traumatology-Medicolegal Aspects (Indian Perspective)akish4uNo ratings yet
- Calcium HydroxideDocument5 pagesCalcium HydroxideCah YaniNo ratings yet
- Target - Online Exam - Oral Pathology Medicine RadiologyDocument58 pagesTarget - Online Exam - Oral Pathology Medicine RadiologyplsssssNo ratings yet
- PhilHealth Circular No. 0035, s.2013 Annex 5 List of Medical Case Rates For Primary Care FacilitiesDocument22 pagesPhilHealth Circular No. 0035, s.2013 Annex 5 List of Medical Case Rates For Primary Care FacilitiesChrysanthus HerreraNo ratings yet
- Failures in Fixed Partial Denture: Presented By: Dr. Jehan Dordi 2 Yr. MdsDocument102 pagesFailures in Fixed Partial Denture: Presented By: Dr. Jehan Dordi 2 Yr. MdsAkanksha MahajanNo ratings yet
- Calcium Hydroxide: A Review: Alireza Farhad and Zahed MohammadiDocument9 pagesCalcium Hydroxide: A Review: Alireza Farhad and Zahed MohammadiÁbilson FabianiNo ratings yet
- Case Discussion 20-9-18Document25 pagesCase Discussion 20-9-18Sarath sasyacNo ratings yet
- Oral Pathology Revision 2010: By: Dr. Sahar El SheikhDocument211 pagesOral Pathology Revision 2010: By: Dr. Sahar El SheikhNada MohammedNo ratings yet
- Clinic Con EndoDocument3 pagesClinic Con EndoVanessa Rose R. FojaNo ratings yet
- DentalDocument42 pagesDentalmahesardarNo ratings yet
- Inter Relation Between Periodontic and The Other Subjects of Dentistry PDFDocument9 pagesInter Relation Between Periodontic and The Other Subjects of Dentistry PDFZakria Al-HadadNo ratings yet
- KODE ICd 10 GIGIDocument2 pagesKODE ICd 10 GIGITio Melda HutagalungNo ratings yet
- Important Tables of Oral PathologyDocument17 pagesImportant Tables of Oral PathologyEasy Med LecturesNo ratings yet
- Periapical GranulomaDocument6 pagesPeriapical GranulomaEzza RiezaNo ratings yet
- Lec 6 EndoDocument14 pagesLec 6 EndoMohamed NabilNo ratings yet
- Effect of Injectable-Platelet Rich Fibrin On MarginalDocument11 pagesEffect of Injectable-Platelet Rich Fibrin On MarginalNurul Fajriah YunitaNo ratings yet
- Q's For CONS VivaDocument251 pagesQ's For CONS VivaHanin AbukhiaraNo ratings yet
- Pulp TherapyDocument8 pagesPulp TherapyKrisna WijayantiNo ratings yet
- Vitapex PDFDocument52 pagesVitapex PDFnadiahapsariNo ratings yet
- Propedeutica EnglDocument7 pagesPropedeutica EnglVicka RainNo ratings yet