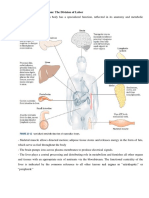
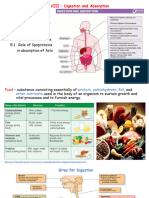
You might also like
- Biochemistry Lectures: Concept of MetabolismDocument4 pagesBiochemistry Lectures: Concept of MetabolismYoussef LatashNo ratings yet
- Intergrated MetabolismDocument117 pagesIntergrated MetabolismDerick SemNo ratings yet
- Introduction To MetabolismDocument33 pagesIntroduction To Metabolismlovelots123488% (16)
- Biochemistry Metabolic Pathways OverviewDocument32 pagesBiochemistry Metabolic Pathways OverviewSanjana VasistNo ratings yet
- Carbohydrates Metabolism NotesDocument15 pagesCarbohydrates Metabolism NotesShaheenNo ratings yet
- Carbohydratemetabolism 140214034339 Phpapp01 ConvertedDocument93 pagesCarbohydratemetabolism 140214034339 Phpapp01 Convertedyixecix709No ratings yet
- Tissue MetabolismDocument13 pagesTissue MetabolismTonny MutisyaNo ratings yet
- Interrelationships of Metabolic PathwaysDocument19 pagesInterrelationships of Metabolic PathwaysAchmad IjaziNo ratings yet
- CARBOHYDRATE METABOLISM-DrSangitaDocument34 pagesCARBOHYDRATE METABOLISM-DrSangitaA MINOR100% (1)
- Metabolism OverviewDocument43 pagesMetabolism OverviewWinnet MaruzaNo ratings yet
- Carbohydrates MetabolismDocument47 pagesCarbohydrates MetabolismDaffa OfficialNo ratings yet
- Carbohydrate Metabolism (Fundamental) Part 1Document20 pagesCarbohydrate Metabolism (Fundamental) Part 1InshaNo ratings yet
- Carbohydrate Metabolism: By:-Dr - Priyanka Sharma 1 Year MDS Dept. of Public Health DentistryDocument93 pagesCarbohydrate Metabolism: By:-Dr - Priyanka Sharma 1 Year MDS Dept. of Public Health DentistrySimham Venu0% (1)
- Metabolism pathways explainedDocument145 pagesMetabolism pathways explainedHesbone AneneNo ratings yet
- Metabolism & Glycolysis PAL - Raghda AdwanDocument74 pagesMetabolism & Glycolysis PAL - Raghda Adwanqueenmasa191No ratings yet
- SEM VI BOTANY C13T Plant MetabolismDocument10 pagesSEM VI BOTANY C13T Plant MetabolismAtashi MandalNo ratings yet
- Cell MetabolismDocument10 pagesCell Metabolismbenishgulzar50No ratings yet
- Lecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyDocument80 pagesLecture 03. Carbohydrate Metabolism. Common Pathway, Regulation and PathologyВіталій Михайлович НечипорукNo ratings yet
- Lecture 12. Liver Biochemistry. Pigmentary Metabolism. JaundicesDocument43 pagesLecture 12. Liver Biochemistry. Pigmentary Metabolism. JaundicesВіталій Михайлович НечипорукNo ratings yet
- Biochemistry of Liver: Alice SkoumalováDocument37 pagesBiochemistry of Liver: Alice Skoumalováandreas kevinNo ratings yet
- Sugar MetabolismDocument113 pagesSugar MetabolismmismaelNo ratings yet
- Chemistry of CarbohydratesDocument10 pagesChemistry of CarbohydratesYahya Daham Zafeer SakhrNo ratings yet
- 2-Energy and RespirationDocument34 pages2-Energy and RespirationLisa DentonNo ratings yet
- Unit 8 Digestion and AbsorptionDocument33 pagesUnit 8 Digestion and AbsorptionGlory MuedaNo ratings yet
- HW - Carbohydrate Metabolism II & Lipid MetabolismDocument2 pagesHW - Carbohydrate Metabolism II & Lipid MetabolismyanNo ratings yet
- Integration of MetabolismDocument17 pagesIntegration of MetabolismSharif M Mizanur RahmanNo ratings yet
- Overview of Metabolism & The Provision of Metabolic Fuels (CHP 16 Harper) - TJLDocument6 pagesOverview of Metabolism & The Provision of Metabolic Fuels (CHP 16 Harper) - TJLMNo ratings yet
- Metabolic Interrelationships Rev 07-11-2014Document82 pagesMetabolic Interrelationships Rev 07-11-2014algutNo ratings yet
- CarbohydratemetabolismDocument88 pagesCarbohydratemetabolismEthio funNo ratings yet
- 16 HF 114 Introduction To MetabolismDocument26 pages16 HF 114 Introduction To Metabolismqueenmasa191No ratings yet
- Metabolism IntegrationDocument10 pagesMetabolism IntegrationValine Cysteine MethionineNo ratings yet
- Stages of Metabolism IDocument29 pagesStages of Metabolism IKiran ShresthaNo ratings yet
- Biochem CH 27 Integration of MetabolismDocument6 pagesBiochem CH 27 Integration of MetabolismSchat ZiNo ratings yet
- CHP 5 Major Metabolic PathwaysDocument53 pagesCHP 5 Major Metabolic PathwaysAmrNo ratings yet
- Guc 2705 59 28561 2023-05-18T10 45 27Document47 pagesGuc 2705 59 28561 2023-05-18T10 45 27malak00mohamed229No ratings yet
- Topic 3 How cells make energyDocument35 pagesTopic 3 How cells make energytmg.35566No ratings yet
- MODULE - FINALS - Biochem - 1Document5 pagesMODULE - FINALS - Biochem - 1Cyril CauilanNo ratings yet
- Carbohydrates Part 2 - MetabolismsDocument60 pagesCarbohydrates Part 2 - MetabolismsNikka Mia AbadiesNo ratings yet
- Carbohydrate MetabolismDocument19 pagesCarbohydrate MetabolismAbhithNo ratings yet
- BiochemieDocument10 pagesBiochemiearian.boostani2002No ratings yet
- Biochemistry - Protein and LipidsDocument31 pagesBiochemistry - Protein and LipidsGiorgi TamazashviliNo ratings yet
- Cellular Metabolism Overview of MetabolismDocument4 pagesCellular Metabolism Overview of MetabolismNathan Louis PalacioNo ratings yet
- Proteins and Amino Acids in BiochemistryDocument8 pagesProteins and Amino Acids in BiochemistryrajanOrajanNo ratings yet
- Mid Biochem: 7. Metabolism Is The Reactions Occurring in A Living Sysstem That Produce and Consume The EnergyDocument2 pagesMid Biochem: 7. Metabolism Is The Reactions Occurring in A Living Sysstem That Produce and Consume The EnergyNguyễn Bá Đạt100% (1)
- Carbohydrate Distribution, Metabolism, and ExcretionDocument93 pagesCarbohydrate Distribution, Metabolism, and ExcretionPSPD - Hana Athaya NNo ratings yet
- Unit-Iv Metabolism 6 Hours Max. Marks: 14 - 16Document36 pagesUnit-Iv Metabolism 6 Hours Max. Marks: 14 - 16Dr. Dhondiba VishwanathNo ratings yet
- CarbohydratesDocument19 pagesCarbohydratesNaina YadavvNo ratings yet
- Integration of Metabolism PathwaysDocument68 pagesIntegration of Metabolism PathwaysCahyani Tiara Safitri100% (1)
- Muscle Energy Metabolsm For MedicineDocument69 pagesMuscle Energy Metabolsm For MedicinefikaduNo ratings yet
- ATP synthesis via two systems in mitochondriaDocument7 pagesATP synthesis via two systems in mitochondriaXuân ViNo ratings yet
- Bioenergetics & Metabolism GuideDocument56 pagesBioenergetics & Metabolism GuideAww AddNo ratings yet
- The Human Metabolism ProcessDocument40 pagesThe Human Metabolism ProcessNur HolikNo ratings yet
- UntitledDocument14 pagesUntitledSyafrianda PanggabeanNo ratings yet
- Carbohydrate Metabolism PathwaysDocument62 pagesCarbohydrate Metabolism PathwaysJHINUK DASGUPTANo ratings yet
- CH 19 Integration of MetabolismDocument63 pagesCH 19 Integration of Metabolismshee140802100% (1)
- Carbohydrate digestion and glucose metabolism pathwaysDocument4 pagesCarbohydrate digestion and glucose metabolism pathwaysJeremy EvansNo ratings yet
- The Role of Liver in Metabolism An Updated ReviewDocument6 pagesThe Role of Liver in Metabolism An Updated Reviewرشاد خالد عماد Rashad Khaled EmadNo ratings yet
- Metabolic Roles of Major Biochemical MoleculesDocument13 pagesMetabolic Roles of Major Biochemical MoleculesJames SoeNo ratings yet
- STM - Merck Case AnswersDocument2 pagesSTM - Merck Case AnswersreetayanNo ratings yet
- Manoshe Street Takeaway MenuDocument9 pagesManoshe Street Takeaway MenuimaddakrNo ratings yet
- IRC Girl Shine Part 4 DigitalDocument124 pagesIRC Girl Shine Part 4 DigitalMónica SalesNo ratings yet
- Shrimp ReceipeDocument2 pagesShrimp Receipedonna blazoNo ratings yet
- Appointment Letter Staff Nurse Job DescriptionDocument3 pagesAppointment Letter Staff Nurse Job DescriptionRenita ChrisNo ratings yet
- RN and LPN Scope of Practice GuideDocument6 pagesRN and LPN Scope of Practice Guidejenn1722No ratings yet
- General Conditions of Contracts For NLNG 2018Document28 pagesGeneral Conditions of Contracts For NLNG 2018AF Dowell MirinNo ratings yet
- Johnson Et Al.2006. Felidae PhylogenyDocument6 pagesJohnson Et Al.2006. Felidae Phylogenyaspergillus_jallNo ratings yet
- Project Execution ProcedureDocument4 pagesProject Execution Procedurecyno2167% (3)
- Jane EyreDocument165 pagesJane EyreSana AbdulsalamNo ratings yet
- InfraTapp - Infra Tapp - Manual ENGDocument64 pagesInfraTapp - Infra Tapp - Manual ENGEliaNo ratings yet
- Production Chemistry and Flow Assurance SimulationDocument21 pagesProduction Chemistry and Flow Assurance SimulationWael Badri100% (1)
- NCM 106 Acute Biologic CrisisDocument142 pagesNCM 106 Acute Biologic CrisisEllamae Chua88% (8)
- FarkolDocument7 pagesFarkolHasiadin LaodeNo ratings yet
- Chap 24 Econ 40Document4 pagesChap 24 Econ 40Racel DelacruzNo ratings yet
- Pre-Final Exam in Audit 2-3Document5 pagesPre-Final Exam in Audit 2-3Shr BnNo ratings yet
- Comparing Food with Comparative AdjectivesDocument4 pagesComparing Food with Comparative AdjectivesLuisito GonzalezNo ratings yet
- DILEM 10 G (24VDC) MoellerDocument7 pagesDILEM 10 G (24VDC) MoellerElizabeth RamirezNo ratings yet
- Track A Beast ManualDocument21 pagesTrack A Beast ManualboltojamNo ratings yet
- Types of ProcurementDocument7 pagesTypes of ProcurementrahulNo ratings yet
- Salary Exchange... A Tax Efficient Way To Boost Your Plans For The FutureDocument4 pagesSalary Exchange... A Tax Efficient Way To Boost Your Plans For The FutureStandard Life UKNo ratings yet
- Menu Baru Kopi GandapoeraDocument7 pagesMenu Baru Kopi GandapoeraAlwan AhpNo ratings yet
- TestDocument4 pagesTestCristina BariNo ratings yet
- Providing Shelter and Safety: A Proposed Evacuation and Rehabilitation Center for Masbate CityDocument62 pagesProviding Shelter and Safety: A Proposed Evacuation and Rehabilitation Center for Masbate CityBenjamae MaqueNo ratings yet
- Effectiveness of Breathing Exercise On Reducing Pain Perception and State Anxiety Among Primi ParturientsDocument8 pagesEffectiveness of Breathing Exercise On Reducing Pain Perception and State Anxiety Among Primi ParturientsPutra SpNo ratings yet
- D754te3 VMDocument37 pagesD754te3 VMGurpreet SinghNo ratings yet
- Akmal Resume BaruDocument3 pagesAkmal Resume BaruSham ZaiNo ratings yet
- A4 RelationshipgridDocument32 pagesA4 RelationshipgridjamesNo ratings yet
- Appendix VIII - Civil and Structural Scope of WorkDocument140 pagesAppendix VIII - Civil and Structural Scope of WorkjaganrajNo ratings yet
- 03 - Henkel Benchmarking (Solutions)Document10 pages03 - Henkel Benchmarking (Solutions)/jncjdncjdnNo ratings yet