You might also like
- Electromyography in CNS Disorders: Central EMGFrom EverandElectromyography in CNS Disorders: Central EMGBhagwan T. ShahaniNo ratings yet
- High-Resolution Measurement of Electrically-Evoked Vagus Nerve Activity in The Anesthetized DogDocument10 pagesHigh-Resolution Measurement of Electrically-Evoked Vagus Nerve Activity in The Anesthetized Dogsuper_montyNo ratings yet
- Norepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaDocument12 pagesNorepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaMariann DuzzNo ratings yet
- Phase Amplitude CouplingDocument8 pagesPhase Amplitude CouplingFideoPereNo ratings yet
- Acupuncture-Stimulated Activation of Sensory NeuronsDocument8 pagesAcupuncture-Stimulated Activation of Sensory NeuronsSam WardNo ratings yet
- The Role of Dorsal Columns Pathway in Visceral Pain: J. PalečekDocument6 pagesThe Role of Dorsal Columns Pathway in Visceral Pain: J. PalečekJorge OchoaNo ratings yet
- Ecam2011 159209Document11 pagesEcam2011 159209Sol Instituto TerapêuticoNo ratings yet
- Nitsche 2004Document5 pagesNitsche 2004Larissa OliveiraNo ratings yet
- Acupuncture Stimulates The Release of Serotonin, But Not Dopamine, in The Rat Nucleus AccumbensDocument6 pagesAcupuncture Stimulates The Release of Serotonin, But Not Dopamine, in The Rat Nucleus Accumbensmarcus_irishNo ratings yet
- Analgesia y ElectroacupunturaDocument8 pagesAnalgesia y ElectroacupunturaagustinNo ratings yet
- Efficient Generation of Reciprocal Signals by Inhibition: K.khodakhah@einstein - Yu.eduDocument42 pagesEfficient Generation of Reciprocal Signals by Inhibition: K.khodakhah@einstein - Yu.eduDulce AguirreNo ratings yet
- Kerrigan 2004Document9 pagesKerrigan 2004Estereotaxia BrasilNo ratings yet
- Megneto TherapyDocument15 pagesMegneto TherapyedcanalNo ratings yet
- Impact of Network Activity On The Integrative PropDocument18 pagesImpact of Network Activity On The Integrative Propttqnhu.rhmNo ratings yet
- Graded Activation of The Intrinsic Laryngeal Muscles For Vocal Fold PosturingDocument7 pagesGraded Activation of The Intrinsic Laryngeal Muscles For Vocal Fold PosturingDaniel AlonsoNo ratings yet
- 5990 FullDocument10 pages5990 FullRomina MendezBrandanNo ratings yet
- K. Morita, R.A. North and T. Tokimasa - Muscarinic Presynaptic Inhibition of Synaptic Transmission in Myenteric Plexus of Guinea-Pig IleumDocument9 pagesK. Morita, R.A. North and T. Tokimasa - Muscarinic Presynaptic Inhibition of Synaptic Transmission in Myenteric Plexus of Guinea-Pig IleumFedrmNo ratings yet
- Bear&KirkwoodDocument11 pagesBear&KirkwoodswagatarcNo ratings yet
- 2802 FullDocument9 pages2802 FullNatalia NuñezNo ratings yet
- Activation of Medial Prefrontal Cortex by Phencyclidine Is Mediated Via A Hippocampo-Prefrontal PathwayDocument7 pagesActivation of Medial Prefrontal Cortex by Phencyclidine Is Mediated Via A Hippocampo-Prefrontal PathwayCortate15gNo ratings yet
- 2007 Transient Modulation of Intracortical InhibitionDocument11 pages2007 Transient Modulation of Intracortical InhibitionchooNo ratings yet
- Kishima 2007Document6 pagesKishima 2007moiNo ratings yet
- NIH Public Access: Author ManuscriptDocument13 pagesNIH Public Access: Author ManuscriptRoar SyltebøNo ratings yet
- 1983 - Partial Infraorbital Nerve Ligation As A Model of Trigeminal Nerve Injury in The Mouse Behavioral, NDocument13 pages1983 - Partial Infraorbital Nerve Ligation As A Model of Trigeminal Nerve Injury in The Mouse Behavioral, NNarawut Paka7eprotNo ratings yet
- Transcallosally Mediated InhibitionDocument11 pagesTranscallosally Mediated InhibitiongiacomopinaliNo ratings yet
- Hypothalamus and Amygdala Response To Acupuncture StimuliDocument13 pagesHypothalamus and Amygdala Response To Acupuncture StimuliAnibal CisternasNo ratings yet
- Akinesia in Parkinsons Disease 1 Shortening of Simple Reaction Time W Focal Single Pulse TMSDocument10 pagesAkinesia in Parkinsons Disease 1 Shortening of Simple Reaction Time W Focal Single Pulse TMSadit020586No ratings yet
- Edwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitsDocument16 pagesEdwards - Neck Muscle Afferents Influence Oromotor and Cardiorespiratory CircuitssbachorickNo ratings yet
- Analgesic Action of Acupuncture and Moxibustion - A Review of Unique Approaches in Japan - 2009.Document7 pagesAnalgesic Action of Acupuncture and Moxibustion - A Review of Unique Approaches in Japan - 2009.alsadicNo ratings yet
- P-Stim™ Auricular Electroacupuncture Stimulation Device For Pain ReliefDocument10 pagesP-Stim™ Auricular Electroacupuncture Stimulation Device For Pain ReliefoanamemoNo ratings yet
- Watanabe 2001Document7 pagesWatanabe 2001Adarsh AbbaganiNo ratings yet
- Knardahl 1998Document7 pagesKnardahl 1998Nurul AzizahNo ratings yet
- Ultrasound Therapy CTADocument5 pagesUltrasound Therapy CTAmitchNo ratings yet
- NITSCHE &PAULUS Excitability Changes Induced in The Human Motor Cortex by Weak Transcranial Direct Current StimulationDocument7 pagesNITSCHE &PAULUS Excitability Changes Induced in The Human Motor Cortex by Weak Transcranial Direct Current Stimulationexcelencia5No ratings yet
- Changes of Extremity and Laryngeal Muscle Electromyographic Amplitudes After Intravenous Administration of Vecuronium00005537-200812000-00014Document5 pagesChanges of Extremity and Laryngeal Muscle Electromyographic Amplitudes After Intravenous Administration of Vecuronium00005537-200812000-00014dr.hungsonNo ratings yet
- Download The Role Of The Dorsal Anterior Insula In Ecstatic Sensation Revealed By Direct Electrical Brain Stimulation F Bartolomei S Lagarde D Scavarda R Carron C G Benar F Picard full chapterDocument42 pagesDownload The Role Of The Dorsal Anterior Insula In Ecstatic Sensation Revealed By Direct Electrical Brain Stimulation F Bartolomei S Lagarde D Scavarda R Carron C G Benar F Picard full chaptersally.rogers984100% (6)
- JNR 23883Document6 pagesJNR 238832021207549No ratings yet
- Paolo Calabresi Et Al - Muscarinic IPSPs in Rat Striatal Cholinergic InterneuronesDocument7 pagesPaolo Calabresi Et Al - Muscarinic IPSPs in Rat Striatal Cholinergic InterneuronesFedrmNo ratings yet
- Baba & Ji (MCN, 2003)Document13 pagesBaba & Ji (MCN, 2003)Jean Pierre Chastre LuzaNo ratings yet
- Van WelieJ Physiol (Lond) 2006Document12 pagesVan WelieJ Physiol (Lond) 2006akrab0101No ratings yet
- C2-Isquemia CardiacaDocument10 pagesC2-Isquemia CardiacaMarta Serra EstopàNo ratings yet
- J Neurophysiol 2000 Ghai 274 80Document8 pagesJ Neurophysiol 2000 Ghai 274 80fnmendoncaNo ratings yet
- 8 5hamaguchiDocument6 pages8 5hamaguchiVhyna Poetri Boegist BoneNo ratings yet
- PNAS 2010 Breshears 1011949107Document6 pagesPNAS 2010 Breshears 1011949107vlmwarrenNo ratings yet
- Abstracts / Brain Stimulation 10 (2017) 346 E540 396Document1 pageAbstracts / Brain Stimulation 10 (2017) 346 E540 396Aleja ToPaNo ratings yet
- Luis Ulloa Nerve StimulationDocument44 pagesLuis Ulloa Nerve StimulationsandraNo ratings yet
- Out 46Document6 pagesOut 46natanatalisNo ratings yet
- Blume 2004Document8 pagesBlume 2004jela1309No ratings yet
- J.F. Cassell & Elspeth M. McLachlan - Muscarinic Agonists Block Five Different Potassium Conductances in Guinea-Pig Sympathetic NeuronesDocument3 pagesJ.F. Cassell & Elspeth M. McLachlan - Muscarinic Agonists Block Five Different Potassium Conductances in Guinea-Pig Sympathetic NeuronesFedrmNo ratings yet
- 10 1016@j Brs 2019 10 022Document49 pages10 1016@j Brs 2019 10 022Alfeku CabraNo ratings yet
- In Vivo Reduction in ATP Cost of Contraction Is Not RelatedDocument11 pagesIn Vivo Reduction in ATP Cost of Contraction Is Not RelatedCharisNo ratings yet
- RAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeDocument14 pagesRAT IN: Bicuculline Potential (EPSP) - Relationship EPSP Amplitude AmplitudeFedrmNo ratings yet
- Chapter 2:: Function Follows Form 29Document3 pagesChapter 2:: Function Follows Form 29Trung Hiệp ĐàoNo ratings yet
- Ijbms 19 800Document4 pagesIjbms 19 800wineniNo ratings yet
- Ning Wang EnglishDocument5 pagesNing Wang EnglishAngela PagliusoNo ratings yet
- Molecular PainDocument16 pagesMolecular PainYixin TongNo ratings yet
- ketamina . NeurocienciaDocument12 pagesketamina . NeurocienciaJosevjemusse21980No ratings yet
- 297 FullDocument10 pages297 FullGaluhFahmiNo ratings yet
- Acupuncure and ImmunomodulationDocument14 pagesAcupuncure and Immunomodulationcristinacaetano9355No ratings yet
- Sleep and Brain Energy LevelsDocument10 pagesSleep and Brain Energy LevelsAngel MartorellNo ratings yet
- Intrinsic Functional Connectivity of The Periaqueductal Gray, A Resting fMRI StudyDocument5 pagesIntrinsic Functional Connectivity of The Periaqueductal Gray, A Resting fMRI StudyYixin TongNo ratings yet
- ForebrainDocument26 pagesForebrainYixin TongNo ratings yet
- Gabbott Et Al-2005-Journal of Comparative NeurologyDocument33 pagesGabbott Et Al-2005-Journal of Comparative NeurologyYixin TongNo ratings yet
- ACC RVM HumanDocument12 pagesACC RVM HumanYixin TongNo ratings yet
- Molecular PainDocument16 pagesMolecular PainYixin TongNo ratings yet
- Lesion of The Rostral Anterior Cingulate Cortex Eliminates The Aversiveness of Spontaneous Neuropathic Pain Following Partial or Complete AxotomyDocument8 pagesLesion of The Rostral Anterior Cingulate Cortex Eliminates The Aversiveness of Spontaneous Neuropathic Pain Following Partial or Complete AxotomyYixin TongNo ratings yet
- Autism Acc v1 FmriDocument12 pagesAutism Acc v1 FmriYixin TongNo ratings yet
- ACC AmygdalaDocument10 pagesACC AmygdalaYixin TongNo ratings yet
- Molecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayDocument20 pagesMolecular Pain: Short-Term Synaptic Plasticity in The Nociceptive Thalamic-Anterior Cingulate PathwayYixin TongNo ratings yet
- Acc SpinalDocument16 pagesAcc SpinalYixin TongNo ratings yet
- Cerebral Hemispheres 2018 PDFDocument69 pagesCerebral Hemispheres 2018 PDFlailatul husnaNo ratings yet
- Physiology of Digestion 2014Document58 pagesPhysiology of Digestion 2014sherumar0% (1)
- Science6 Q2 Mod3 ExplainHowTheDifferentOrganSystemWorksTogether V4Document24 pagesScience6 Q2 Mod3 ExplainHowTheDifferentOrganSystemWorksTogether V4Jonel BuergoNo ratings yet
- Interoceptive Rhythms in The BrainDocument15 pagesInteroceptive Rhythms in The BrainAlisson Araya HNo ratings yet
- Artikel Asli Artikel Asli: Volume 3, Nomor 2 Mei - Agustus 2018Document6 pagesArtikel Asli Artikel Asli: Volume 3, Nomor 2 Mei - Agustus 2018Khotimah AdeNo ratings yet
- 10 - Histology Lecture, Structure of Muscular TissueDocument37 pages10 - Histology Lecture, Structure of Muscular TissueAMIRA HELAYELNo ratings yet
- Terms and Strategies For Medical RecordsDocument42 pagesTerms and Strategies For Medical RecordsJesus ParragaNo ratings yet
- Surgical Group - Normal Physiologic Changes in Aging Affecting Various SystemsDocument20 pagesSurgical Group - Normal Physiologic Changes in Aging Affecting Various SystemsKyra Bianca R. FamacionNo ratings yet
- Gastric Hormones and Pancreatic EnzymesDocument7 pagesGastric Hormones and Pancreatic EnzymesmyscribeNo ratings yet
- 1 ADocument15 pages1 AzuliaahmadNo ratings yet
- Digestive System Workshop For Kids by SlidesgoDocument16 pagesDigestive System Workshop For Kids by Slidesgoapi-649419165No ratings yet
- THE Gastro-Intestinal TractDocument51 pagesTHE Gastro-Intestinal TractandrewNo ratings yet
- Structure and Functions of The Ear Explicated With DiagramsDocument10 pagesStructure and Functions of The Ear Explicated With DiagramsemmanuelNo ratings yet
- 7 The BrainDocument19 pages7 The BrainZidane ZizouNo ratings yet
- Kasus 1 Tiroid-1Document18 pagesKasus 1 Tiroid-1Ratih Sri rezekiNo ratings yet
- SIADH, DI and Cerebral Salt Wasting: Karim Rafaat, MDDocument54 pagesSIADH, DI and Cerebral Salt Wasting: Karim Rafaat, MDMahendra PrasetyoNo ratings yet
- Essentials of Human PhysiologyDocument552 pagesEssentials of Human PhysiologyTapan Kumar Pal100% (4)
- Circulatory System Gizmo - 5 Mark TestDocument3 pagesCirculatory System Gizmo - 5 Mark TestDhruv VmNo ratings yet
- Essay # 4. DigeDocument9 pagesEssay # 4. DigeAbdul Karim BurkiNo ratings yet
- DLP in Science 4 (7 E) (Kidney)Document3 pagesDLP in Science 4 (7 E) (Kidney)Aiza Tolentino-Carpio100% (2)
- Case Study AGE With Signs of DehydrationDocument27 pagesCase Study AGE With Signs of Dehydrationtansincos100% (23)
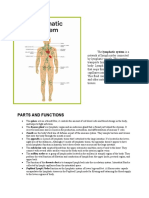
- Parts and FunctionsDocument4 pagesParts and FunctionsGerwin SilvestreNo ratings yet
- Day Study Schedule For USMLE Step 1 LecturioDocument12 pagesDay Study Schedule For USMLE Step 1 LecturioDany Paulbaby100% (1)
- Chapter 12 Conception and Fetal DevelopmentDocument17 pagesChapter 12 Conception and Fetal DevelopmentRoneshia Michelle Rudolph100% (1)
- Hans Selye and The Field of Stress Research A Syndrome Produced by Diverse Nocuous AgentsDocument2 pagesHans Selye and The Field of Stress Research A Syndrome Produced by Diverse Nocuous AgentsMar RajNo ratings yet
- Consumer Behaviour Asia Pacific 1st Edition Hoyer Test BankDocument26 pagesConsumer Behaviour Asia Pacific 1st Edition Hoyer Test BankMatthewMossfnak100% (55)
- Liver CirrhosisDocument30 pagesLiver CirrhosisMelody PardilloNo ratings yet
- Puberty and The Tanner StagesDocument2 pagesPuberty and The Tanner StagesPramedicaPerdanaPutraNo ratings yet
- 181 Anatomy Eyes and EarsDocument23 pages181 Anatomy Eyes and EarsAnita YadavNo ratings yet
- Benign Breast DisordersDocument30 pagesBenign Breast DisordersShams Nadeem AlamNo ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (17)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (403)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- The Ultimate Guide To Memory Improvement TechniquesFrom EverandThe Ultimate Guide To Memory Improvement TechniquesRating: 5 out of 5 stars5/5 (34)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- Techniques Exercises And Tricks For Memory ImprovementFrom EverandTechniques Exercises And Tricks For Memory ImprovementRating: 4.5 out of 5 stars4.5/5 (40)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Algorithms to Live By: The Computer Science of Human DecisionsFrom EverandAlgorithms to Live By: The Computer Science of Human DecisionsRating: 4.5 out of 5 stars4.5/5 (722)
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)