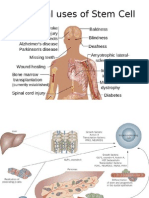
You might also like
- DR Schulze BlogoviskDocument241 pagesDR Schulze BlogoviskMitarMitrovic100% (4)
- Malunion Delayed Union and Nonunion FracturesDocument31 pagesMalunion Delayed Union and Nonunion FracturesAzmi FarhadiNo ratings yet
- Complete Head To Toe AssessmentDocument7 pagesComplete Head To Toe AssessmentClyde100% (1)
- Isolation and Characterisation of Mesenchymal Stem Cells From Adult Mouse Bone MarrowDocument12 pagesIsolation and Characterisation of Mesenchymal Stem Cells From Adult Mouse Bone MarrowRamasamy19No ratings yet
- ArthritsDocument27 pagesArthritsLakshmipriya GopinathNo ratings yet
- Pre ResechDocument20 pagesPre ResechWida MarianeNo ratings yet
- Braid Et Al 2018Document13 pagesBraid Et Al 2018Cahaya Carla BangsawanNo ratings yet
- Research Article in Vitro and in VivoDocument14 pagesResearch Article in Vitro and in VivoVILEOLAGOLDNo ratings yet
- Pluripotent Stem Cell BiologyDocument227 pagesPluripotent Stem Cell BiologyhrbasuNo ratings yet
- Wang 2009Document13 pagesWang 2009Novi ArthaNo ratings yet
- Stem Cell Therapy: Jessica M. QuimbyDocument9 pagesStem Cell Therapy: Jessica M. QuimbySJ JungNo ratings yet
- Safety Study of Allogeneic Mesenchymal Stem Cell Therapy in Animal-MainDocument8 pagesSafety Study of Allogeneic Mesenchymal Stem Cell Therapy in Animal-Mainsitiaminah.muhammadimranNo ratings yet
- Bahan Buat Presentasi Stem Cell DMDocument11 pagesBahan Buat Presentasi Stem Cell DMArfan PrawiragaraNo ratings yet
- Comparing The Therapeutic Potential of Stem CellsDocument30 pagesComparing The Therapeutic Potential of Stem CellsAnaaya SinghaniaNo ratings yet
- Strategies To Capitalize On Cell Spheroid Therapeutic Potential For Tissue Repair and Disease ModelingDocument13 pagesStrategies To Capitalize On Cell Spheroid Therapeutic Potential For Tissue Repair and Disease ModelingShai FuchsNo ratings yet
- ღეროვანი უჯრედებიDocument10 pagesღეროვანი უჯრედებიIrakli GhlontiNo ratings yet
- Haem 2007 cxcr4 MSC HomingDocument8 pagesHaem 2007 cxcr4 MSC HomingoaifoiweuNo ratings yet
- Equine Yolk Sac: A Stem Cells Source: Saco Vitelino Equino: Una Fuente de Células MadreDocument9 pagesEquine Yolk Sac: A Stem Cells Source: Saco Vitelino Equino: Una Fuente de Células MadreisabelNo ratings yet
- tmp3831 TMPDocument3 pagestmp3831 TMPFrontiersNo ratings yet
- Jurnal KJH-Derivation of Mesenchymal Stem Cells From HumanDocument9 pagesJurnal KJH-Derivation of Mesenchymal Stem Cells From HumanClarion 642No ratings yet
- Cell Biology:: Extracellular Supportive and Unsupportive Comparative Proteomic Analysis ofDocument17 pagesCell Biology:: Extracellular Supportive and Unsupportive Comparative Proteomic Analysis ofGeasle DerremenNo ratings yet
- Cap 5Document14 pagesCap 5NatsumiTakamoriNo ratings yet
- 2017 Article 199Document13 pages2017 Article 199Rabiatul 'raney' AdawiyahNo ratings yet
- Human Adipose-Derived Stem Cells Inhibit Bioactivity of Keloid FibroblastsDocument8 pagesHuman Adipose-Derived Stem Cells Inhibit Bioactivity of Keloid FibroblastssesiaNo ratings yet
- Xin 2013Document5 pagesXin 2013Maximiliano PfannlNo ratings yet
- Braga C Cytotherapy 2022 Proteomics of MSC EV Normoxia HypoxiaDocument14 pagesBraga C Cytotherapy 2022 Proteomics of MSC EV Normoxia HypoxiaAlmira AlodiaNo ratings yet
- Bone Marrow Mesenchymal Stem Cells For Improving Hematopoietic Function An in Vitro and in Vivo Model On Bone Marrow MicroenvironmentDocument9 pagesBone Marrow Mesenchymal Stem Cells For Improving Hematopoietic Function An in Vitro and in Vivo Model On Bone Marrow Microenvironmentyanyan wangNo ratings yet
- Author's Accepted Manuscript: Seminars in Pediatric SurgeryDocument28 pagesAuthor's Accepted Manuscript: Seminars in Pediatric Surgerywibm23No ratings yet
- Stirred Suspension Bioreactor Culture of Porcine Induced Pluripotent Stem CellsDocument12 pagesStirred Suspension Bioreactor Culture of Porcine Induced Pluripotent Stem CellsArianna CastroNo ratings yet
- Mesenchymal Stem Cells Literature ReviewDocument9 pagesMesenchymal Stem Cells Literature ReviewafdtygyhkNo ratings yet
- JMCC2003 YoshidaDocument23 pagesJMCC2003 Yoshidaslmkyl1805No ratings yet
- International ImmunopharmacologyDocument15 pagesInternational ImmunopharmacologyAndres RamirezNo ratings yet
- 2022 International Immunopharmacology Q1RamírezDocument15 pages2022 International Immunopharmacology Q1RamírezAndres RamirezNo ratings yet
- Stem Cell Secretome and Its EffectsDocument34 pagesStem Cell Secretome and Its EffectsMaría del Rosario Tudela MuleroNo ratings yet
- Original Article: Bhaskar Vyas, Atul Shah, Anant Marathe, Ansarullah, Rajni Vyas, Ramesh BhondeDocument5 pagesOriginal Article: Bhaskar Vyas, Atul Shah, Anant Marathe, Ansarullah, Rajni Vyas, Ramesh BhondeAtul ShahNo ratings yet
- 1 s2.0 S018844092030638X MainDocument9 pages1 s2.0 S018844092030638X MainDarshan GandhiNo ratings yet
- 2012 Article 9528Document9 pages2012 Article 9528Lê Thùy NgânNo ratings yet
- STEM CELLS - 2009 - Kusumbe - CD133 Expressing Stem Cells Associated With Ovarian Metastases Establish An EndothelialDocument11 pagesSTEM CELLS - 2009 - Kusumbe - CD133 Expressing Stem Cells Associated With Ovarian Metastases Establish An EndothelialJyoti DeshmukhNo ratings yet
- SCD 2009 0138Document16 pagesSCD 2009 0138Gonçalo NunesNo ratings yet
- Cells: Fferences Between The Proliferative Effects ofDocument12 pagesCells: Fferences Between The Proliferative Effects ofmeltwithsnow163.comNo ratings yet
- Stemcells 2007-0416Document12 pagesStemcells 2007-0416Ikke OppgaveNo ratings yet
- AnimalCellCulture Eng TSDocument12 pagesAnimalCellCulture Eng TSsumitadas4477No ratings yet
- tmpDBD5 TMPDocument6 pagestmpDBD5 TMPFrontiersNo ratings yet
- Lab 2-4 Formal ReportDocument7 pagesLab 2-4 Formal ReportAbdullah AslamNo ratings yet
- Ncomms12015 Scleroza Multipla PDFDocument11 pagesNcomms12015 Scleroza Multipla PDFLUTICHIEVICI NATALIANo ratings yet
- Yarani R Cytotherapy 2023 MSC-EVDocument2 pagesYarani R Cytotherapy 2023 MSC-EVAlmira AlodiaNo ratings yet
- S74 Poster Abstracts: Table 1Document1 pageS74 Poster Abstracts: Table 1HanaNo ratings yet
- The Spatial Transcriptomic Landscape of The Healing Mouse Intestine Following DamageDocument16 pagesThe Spatial Transcriptomic Landscape of The Healing Mouse Intestine Following Damageyjbu0125No ratings yet
- s12014 020 09279 6Document18 pagess12014 020 09279 6laryss.csantosNo ratings yet
- Breast Cancer Stem Cells Derived Extracellular Vesicles Affe - 2022 - Clinical BDocument13 pagesBreast Cancer Stem Cells Derived Extracellular Vesicles Affe - 2022 - Clinical BAd AdrianaNo ratings yet
- Extracellular Vesicles (Ber)Document13 pagesExtracellular Vesicles (Ber)Farhan Royan PermanahadiNo ratings yet
- 1 s2.0 S1465324917300051 MainDocument15 pages1 s2.0 S1465324917300051 MainPando RaitaNo ratings yet
- JCR 23 002Document18 pagesJCR 23 002yakalapunagarajuNo ratings yet
- MSC Reduce IL-6 NewDocument12 pagesMSC Reduce IL-6 NewFarhan Royan PermanahadiNo ratings yet
- Exosomes Derived From Mir-26A-Modified Mscs Promote Axonal Regeneration Via The Pten/Akt/Mtor Pathway Following Spinal Cord InjuryDocument15 pagesExosomes Derived From Mir-26A-Modified Mscs Promote Axonal Regeneration Via The Pten/Akt/Mtor Pathway Following Spinal Cord Injuryelif.onsozNo ratings yet
- Cells: Fferential Regulation of Circrna, Mirna, and Fferentiation of Human Mesenchymal Stromal CellsDocument24 pagesCells: Fferential Regulation of Circrna, Mirna, and Fferentiation of Human Mesenchymal Stromal CellsBreeze ChloeNo ratings yet
- 1 อนุภาคนาโนที่ได้จากโสมจะเปลี่ยนขั้วของมDocument18 pages1 อนุภาคนาโนที่ได้จากโสมจะเปลี่ยนขั้วของมTheppitak TheppitakNo ratings yet
- Adipose Derived Stem Migration in The Vascular System After Transplantation and The Potential Colonization of Ectopic Sites in SwineDocument21 pagesAdipose Derived Stem Migration in The Vascular System After Transplantation and The Potential Colonization of Ectopic Sites in SwineAthenaeum Scientific PublishersNo ratings yet
- Chemically Induced Reprogramming of Somatic Cells To Pluripotent Stem Cells and Neural CellsDocument13 pagesChemically Induced Reprogramming of Somatic Cells To Pluripotent Stem Cells and Neural CellsnembutalNo ratings yet
- Research Article: Cellular Response To A Novel Fetal Acellular Collagen Matrix: Implications For Tissue RegenerationDocument9 pagesResearch Article: Cellular Response To A Novel Fetal Acellular Collagen Matrix: Implications For Tissue Regenerationa27376181No ratings yet
- Ha Et Al. - 2021 - Human Dental Pulp-Derived Mesenchymal Stem Cell PoDocument13 pagesHa Et Al. - 2021 - Human Dental Pulp-Derived Mesenchymal Stem Cell PoEbaycustomer number01No ratings yet
- Biomaterials That Promote Cell-Cell Interactions Enhance The Paracrine Function of MSCsDocument12 pagesBiomaterials That Promote Cell-Cell Interactions Enhance The Paracrine Function of MSCsSadie LaniganNo ratings yet
- Agricultural InjuryDocument21 pagesAgricultural InjuryAzmi FarhadiNo ratings yet
- NexusDocument6 pagesNexusAzmi FarhadiNo ratings yet
- Subaidah 7/1/1958 362516 Ods Glaukoma Os Pe + Iol FD WA GA 2Document1 pageSubaidah 7/1/1958 362516 Ods Glaukoma Os Pe + Iol FD WA GA 2Azmi FarhadiNo ratings yet
- Revised - IJHES - The Existence and Management of Islamic Boarding SchoolsDocument17 pagesRevised - IJHES - The Existence and Management of Islamic Boarding SchoolsAzmi FarhadiNo ratings yet
- Acute Compartment Syndrome in Lower Extremity JAAOSDocument9 pagesAcute Compartment Syndrome in Lower Extremity JAAOSAzmi FarhadiNo ratings yet
- Marfan Syndrome A Clinical Update JAAOSDocument7 pagesMarfan Syndrome A Clinical Update JAAOSAzmi FarhadiNo ratings yet
- Damage Control Orthopaedics in Spinal Trauma JAAOSDocument12 pagesDamage Control Orthopaedics in Spinal Trauma JAAOSAzmi FarhadiNo ratings yet
- Subaxial Cervical Spine Trauma JAAOSDocument12 pagesSubaxial Cervical Spine Trauma JAAOSAzmi FarhadiNo ratings yet
- Hannover Polytrauma Score.4Document2 pagesHannover Polytrauma Score.4Azmi FarhadiNo ratings yet
- Venous Thromboembolism in MalignancyDocument3 pagesVenous Thromboembolism in MalignancyAzmi FarhadiNo ratings yet
- Extensor Tendon Injuries in The Hand JAAOSDocument8 pagesExtensor Tendon Injuries in The Hand JAAOSAzmi FarhadiNo ratings yet
- Primary Osteoporosis in Postmenopausal WomenDocument5 pagesPrimary Osteoporosis in Postmenopausal WomenAzmi FarhadiNo ratings yet
- Spondylolysis and Spondylolisthesis in Children and Adolescents JAAOSDocument8 pagesSpondylolysis and Spondylolisthesis in Children and Adolescents JAAOSAzmi FarhadiNo ratings yet
- Varus Osteotomy French 1959Document3 pagesVarus Osteotomy French 1959Azmi FarhadiNo ratings yet
- Safe Tourniquet Use A Review of The Evidence JAAOSDocument10 pagesSafe Tourniquet Use A Review of The Evidence JAAOSAzmi FarhadiNo ratings yet
- Iub 1216Document7 pagesIub 1216Azmi FarhadiNo ratings yet
- Fat EmbolismDocument10 pagesFat EmbolismAzmi FarhadiNo ratings yet
- TB Arthritis Ankle Mimicking SynovitisDocument8 pagesTB Arthritis Ankle Mimicking SynovitisAzmi FarhadiNo ratings yet
- Supracondylar Fractures of The Humerus in Children.3Document8 pagesSupracondylar Fractures of The Humerus in Children.3Azmi FarhadiNo ratings yet
- True Ponte Osteotomy Ponte 2018Document10 pagesTrue Ponte Osteotomy Ponte 2018Azmi FarhadiNo ratings yet
- Discuss Tension Band Principles: by DR Kabiru SALISU 11 Aug. 2017Document40 pagesDiscuss Tension Band Principles: by DR Kabiru SALISU 11 Aug. 2017Azmi FarhadiNo ratings yet
- Sural BistaDocument6 pagesSural BistaAzmi FarhadiNo ratings yet
- 18 Rosenwasser Distal Radius Fractures HandoutDocument13 pages18 Rosenwasser Distal Radius Fractures HandoutAzmi FarhadiNo ratings yet
- The Pins and Rubbers Traction System FOR Treatment of Comminuted Intraarticular Fractures and Fracture-Dislocations in The HandDocument10 pagesThe Pins and Rubbers Traction System FOR Treatment of Comminuted Intraarticular Fractures and Fracture-Dislocations in The HandAzmi FarhadiNo ratings yet
- Hand and Wrist ExaminationDocument49 pagesHand and Wrist ExaminationAzmi FarhadiNo ratings yet
- A Sept Abi LitasDocument28 pagesA Sept Abi LitasAzmi FarhadiNo ratings yet
- Field Work Report Sher Zada (Role of DOST Foundation)Document37 pagesField Work Report Sher Zada (Role of DOST Foundation)KHAN_EROS100% (2)
- Hypogammaglobulinemia and ArthritisDocument36 pagesHypogammaglobulinemia and ArthritisNilesh NolkhaNo ratings yet
- Seasonal Affective Disorder StudyDocument7 pagesSeasonal Affective Disorder Studyvidro3No ratings yet
- Hempel's Curing Agent 9525049810 En-UsDocument10 pagesHempel's Curing Agent 9525049810 En-UssasikumareNo ratings yet
- Peter Fragatos 1 SubstanceAbuse MentalCondition PDFDocument41 pagesPeter Fragatos 1 SubstanceAbuse MentalCondition PDFWKYC.comNo ratings yet
- Abdominal AssesssmentDocument113 pagesAbdominal Assesssmentshamy143No ratings yet
- Avian and Exotic Pets - 2009Document310 pagesAvian and Exotic Pets - 2009Rouge07No ratings yet
- The Pathophysiology of Peptic UlcerDocument15 pagesThe Pathophysiology of Peptic UlcerKike Meneses100% (1)
- Physical Therapy - Mike Boyle Strength & ConditioningDocument2 pagesPhysical Therapy - Mike Boyle Strength & Conditioningjsdfskjdhf100% (1)
- AH 2019 Post-Show-ReportDocument11 pagesAH 2019 Post-Show-ReportPermesh GoelNo ratings yet
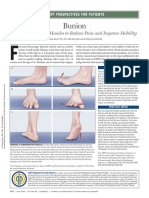
- Bunion: Strengthening Foot Muscles To Reduce Pain and Improve MobilityDocument1 pageBunion: Strengthening Foot Muscles To Reduce Pain and Improve Mobilitymucki_provokatorNo ratings yet
- Curriculum Vitae Dirk J.M.K. de Ridder, M.D., PH.D., FEBUDocument16 pagesCurriculum Vitae Dirk J.M.K. de Ridder, M.D., PH.D., FEBUمحمد عريف حكيمNo ratings yet
- A Digital Inclusion Project For Older PeopleDocument1 pageA Digital Inclusion Project For Older PeopleMauricio AraujoNo ratings yet
- LunaDocument24 pagesLunaapi-471513506No ratings yet
- Suturing SkillsDocument41 pagesSuturing Skillscj barias100% (1)
- Book Forum - On HysteriaDocument15 pagesBook Forum - On Hysteriaadmin1666No ratings yet
- Uso Terapeutico Del Tramadol Como Alternativa Analgesica en El Aguola de HarrisDocument83 pagesUso Terapeutico Del Tramadol Como Alternativa Analgesica en El Aguola de HarrisCamila Rodríguez AvilánNo ratings yet
- Anticoagulant TherapyDocument47 pagesAnticoagulant TherapyAndriKusuma75% (4)
- Who Can Donate BloodDocument4 pagesWho Can Donate BloodsreeramNo ratings yet
- Denture Trouble Shooting GuideDocument5 pagesDenture Trouble Shooting Guideheri asriyadiNo ratings yet
- Hoja Seguridad Dromus BLDocument7 pagesHoja Seguridad Dromus BLClaudia hernandezNo ratings yet
- Lipidocare User GuideDocument89 pagesLipidocare User GuideCarlos Alberto ContrerasqNo ratings yet
- The Effects of Prophylactic Bolus Phenylephrine On Hypotension During Low-Dose Spinal Anesthesia For Cesarean SectionDocument6 pagesThe Effects of Prophylactic Bolus Phenylephrine On Hypotension During Low-Dose Spinal Anesthesia For Cesarean SectionAgus GunardiNo ratings yet
- Product Catalogue 2017 Food and Feed Analysis enDocument82 pagesProduct Catalogue 2017 Food and Feed Analysis enNelson AcostaNo ratings yet
- Infusion Pump & Syringe Pump RevisedDocument18 pagesInfusion Pump & Syringe Pump Revisedfrances182871% (7)
- Microchem ASTM E2315 Study Report NG6838 05MAR2016Document9 pagesMicrochem ASTM E2315 Study Report NG6838 05MAR2016Vilva ManikandanNo ratings yet
- Vol 26.4 - Sleep Neurology.2020Document299 pagesVol 26.4 - Sleep Neurology.2020Mardika FamilyNo ratings yet
- Cis Self-Study Lesson PlanDocument4 pagesCis Self-Study Lesson Planjerimiah_manzonNo ratings yet