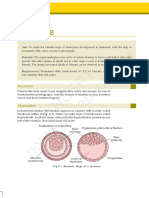
You might also like
- 06 GastrulationtxtDocument38 pages06 GastrulationtxtHafidzul HalimNo ratings yet
- Principles of Genetics and Animal BreedingDocument18 pagesPrinciples of Genetics and Animal BreedingJoshua BadongNo ratings yet
- RegenerationDocument17 pagesRegenerationtiestonNo ratings yet
- Rat Urinary and Reproductive SystemDocument4 pagesRat Urinary and Reproductive SystemAnkit NariyaNo ratings yet
- Frog Embryo Development StagesDocument3 pagesFrog Embryo Development Stagesangel_soshiNo ratings yet
- Animal Form and Function: C E L PDocument38 pagesAnimal Form and Function: C E L PJoshua Miguel Buno0% (1)
- Sex Determination in DrosophilaDocument19 pagesSex Determination in DrosophilaSheerin Sulthana100% (2)
- Cytoplasmic InheritanceDocument13 pagesCytoplasmic InheritanceIzza NafisaNo ratings yet
- Respiratory System: StructureDocument29 pagesRespiratory System: StructureDr. Abir Ishtiaq100% (1)
- Metamorphosis in InsectsDocument13 pagesMetamorphosis in InsectsAnupam GhoshNo ratings yet
- Manual Paper I FinalDocument199 pagesManual Paper I Finalankit pandey100% (1)
- Species Concepts ExplainedDocument26 pagesSpecies Concepts ExplainedmashedtomatoezedNo ratings yet
- Gametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalDocument16 pagesGametogenesis: Spermatogenesis & Oogenesis: Dr. Naveen Kumar MMMC ManipalLim Yi QiNo ratings yet
- Insect Reproduction CK Dash SAUDocument3 pagesInsect Reproduction CK Dash SAUChandra Kanta Dash100% (1)
- Chromosomal sex determination systems overviewDocument61 pagesChromosomal sex determination systems overviewDeepak maliNo ratings yet
- Dynamics of The Mammalian Sperm Plasma Membrane in The Process of FertilizationDocument39 pagesDynamics of The Mammalian Sperm Plasma Membrane in The Process of FertilizationWachiel ArhamzNo ratings yet
- TOrsionDocument19 pagesTOrsionDrSneha VermaNo ratings yet
- Early DevelopmentDocument5 pagesEarly DevelopmentMarina of The SeaNo ratings yet
- Human Reroduction by Digambar Mali.Document22 pagesHuman Reroduction by Digambar Mali.Surge E-Solution Pvt.Ltd.100% (1)
- Torsion in GastropodsDocument3 pagesTorsion in GastropodsHabiba100% (2)
- MSC Semester I E-Content Classification of FungiDocument8 pagesMSC Semester I E-Content Classification of FungiNAYAN JYOTI KALITA100% (1)
- Parasitic Adaptation PDFDocument6 pagesParasitic Adaptation PDFmubarik Hayat50% (2)
- Lecture 1B - Benzer's Work and Complementation TestDocument5 pagesLecture 1B - Benzer's Work and Complementation TestLAKSHAY VIRMANI100% (1)
- Definition of AllelochemicalsDocument7 pagesDefinition of AllelochemicalsrubyNo ratings yet
- Spirogyra: Weed) Is A Genus of Filamentous Chlorophyte Green Algae of The OrderDocument12 pagesSpirogyra: Weed) Is A Genus of Filamentous Chlorophyte Green Algae of The Orderx456456456xNo ratings yet
- CASA Krishnakumar2011Document9 pagesCASA Krishnakumar2011Oriza Savitri AriantieNo ratings yet
- Biopharming SlideDocument19 pagesBiopharming SlideSN ParvezNo ratings yet
- Embryology - Science That Studies The Early Development and Formation of The OrganismDocument27 pagesEmbryology - Science That Studies The Early Development and Formation of The OrganismMeah PachecoNo ratings yet
- Animal Kingdom Non ChordataDocument26 pagesAnimal Kingdom Non ChordataRaichal P BijuNo ratings yet
- Histo NotesDocument19 pagesHisto Notesanas ishaqNo ratings yet
- Protein TargettingDocument11 pagesProtein TargettingPMIB Matrikulasi FKUI 2018/2019No ratings yet
- Developmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Document48 pagesDevelopmental Biology, 12th Edition (Michael J.F. Barresi, Scott F. Gilbert)Nita DelinaNo ratings yet
- Assignment: SubjectDocument4 pagesAssignment: SubjectAbdullah BhattiNo ratings yet
- Carbohydrates: Definition, Structure, Properties and ClassificationDocument45 pagesCarbohydrates: Definition, Structure, Properties and ClassificationKiya Alemu100% (1)
- Biology - Zoology: Practical ManualDocument24 pagesBiology - Zoology: Practical ManualMohammed umar sheriff100% (1)
- Animal Kingdom-Subphylum Vertebrata-Class IXDocument2 pagesAnimal Kingdom-Subphylum Vertebrata-Class IXMonika Mehan100% (2)
- Animal BreedingDocument37 pagesAnimal BreedingRencsNo ratings yet
- ETEA key animal kingdom notesDocument2 pagesETEA key animal kingdom notesUsama Iqbal33% (3)
- Lab Report 4 GametogenesisDocument13 pagesLab Report 4 GametogenesisWnz NaiveNo ratings yet
- Protein Sorting and TransportDocument17 pagesProtein Sorting and TransportMichiko BernardoNo ratings yet
- Nutrition in ProtozoaDocument30 pagesNutrition in ProtozoaDIV TLM ARIFA ALHUSNAH100% (1)
- Lab 10 Porifera & CnidariaDocument25 pagesLab 10 Porifera & Cnidariadraykid100% (1)
- Juyena Et Al-2012-Journal of AndrologyDocument16 pagesJuyena Et Al-2012-Journal of AndrologyAnamaria Blaga PetreanNo ratings yet
- Introduction To Developmental BiologyDocument39 pagesIntroduction To Developmental BiologyBinita SedhaiNo ratings yet
- Chicken Embryo Lab ReportDocument10 pagesChicken Embryo Lab Reportgmk5031No ratings yet
- 2b. Types of Egg and CleavageDocument55 pages2b. Types of Egg and CleavageManisha BishtNo ratings yet
- 1 - Sex DeterminationDocument18 pages1 - Sex Determinationmis_administratorNo ratings yet
- Phylum NematodaDocument24 pagesPhylum NematodaKaten KyoukotsuNo ratings yet
- Organogenesis: By, BIRAC Innovation Fellow University of Agricultural Sciences, DharwadDocument24 pagesOrganogenesis: By, BIRAC Innovation Fellow University of Agricultural Sciences, DharwadTabada NickyNo ratings yet
- General Classification (All Phylum)Document16 pagesGeneral Classification (All Phylum)kiran PoudelNo ratings yet
- Biomolecules: Classification and FunctionsDocument18 pagesBiomolecules: Classification and Functionsdrugdrug100% (1)
- Chordate Characteristics and ClassificationDocument28 pagesChordate Characteristics and ClassificationNil BanerjeeNo ratings yet
- 2006 Cryopreservation and Intracytoplasmic Sperm Injection With Bovine Eith Egg Yolk Sigma E0625Document215 pages2006 Cryopreservation and Intracytoplasmic Sperm Injection With Bovine Eith Egg Yolk Sigma E0625Oriza Savitri AriantieNo ratings yet
- Subject - Zoology B.sc. Part-I (Semester I & II)Document11 pagesSubject - Zoology B.sc. Part-I (Semester I & II)Sanjeev ChaudharyNo ratings yet
- Biology Notes Chap: 3Document5 pagesBiology Notes Chap: 3R.S.HNo ratings yet
- Classification of AnimalDocument12 pagesClassification of AnimalMustika Ayuni100% (1)
- Lsc-Cyfair: Bird MigrationDocument40 pagesLsc-Cyfair: Bird MigrationNaman DhuaanNo ratings yet
- Exercise 7 Chick EmbryoDocument11 pagesExercise 7 Chick Embryokoshi sugawaraNo ratings yet
- Chapter 33 - Descriptive EmbryologyDocument12 pagesChapter 33 - Descriptive Embryologyngquvu67% (15)
- CleavageDocument4 pagesCleavageapi-38010390% (1)
- Iosob 2008 Observations On The Embryonic Devepolment of Triturus Lissotriton Vulgaris LDocument12 pagesIosob 2008 Observations On The Embryonic Devepolment of Triturus Lissotriton Vulgaris LGabrielNo ratings yet
- Book - The Frog Its Reproduction and Development 17 - EmbryologyDocument12 pagesBook - The Frog Its Reproduction and Development 17 - EmbryologyJudy Anne Vitales GalizaNo ratings yet
- SnodgrassDocument513 pagesSnodgrassCelestina Nebulosa100% (3)
- Zoology Multiple Choice Question (GuruKpo)Document33 pagesZoology Multiple Choice Question (GuruKpo)GuruKPO100% (2)
- 04 Laboratory Manual Sbc3043 Semester 1, 2018 2019 A181Document19 pages04 Laboratory Manual Sbc3043 Semester 1, 2018 2019 A181Leena MuniandyNo ratings yet
- © Ncert Not To Be Republished: Exercise 8Document5 pages© Ncert Not To Be Republished: Exercise 8Ankit YadavNo ratings yet
- Comparative Anatomy and Developmental Biology of VertebratesDocument2 pagesComparative Anatomy and Developmental Biology of VertebratesMalik JunaidNo ratings yet
- Embryonic Development Stages from Cleavage to NeurulationDocument41 pagesEmbryonic Development Stages from Cleavage to NeurulationHyenaNo ratings yet
- Chapter 5 Comparative Vertebrate AnatomyDocument11 pagesChapter 5 Comparative Vertebrate AnatomyNathaniel Arthur EstavilloNo ratings yet
- Human Embryonic DevelopmentDocument8 pagesHuman Embryonic DevelopmentGargi BhattacharyaNo ratings yet
- DAT Bootcamp - Developmental Biology Study GuideDocument6 pagesDAT Bootcamp - Developmental Biology Study GuidePearl PascuaNo ratings yet
- КлонированиеDocument2 pagesКлонированиеRed FoxNo ratings yet
- 12 Development TutorialDocument63 pages12 Development TutorialBabasChongNo ratings yet
- Frog Embryology PDFDocument17 pagesFrog Embryology PDFHohai ConcepcionNo ratings yet
- Human Development Prenatal StageDocument2 pagesHuman Development Prenatal StageZie BeaNo ratings yet
- Comparative Embryology, Aspects of Human Reproduction Pollution and Its Effect On OrganismsDocument35 pagesComparative Embryology, Aspects of Human Reproduction Pollution and Its Effect On OrganismsSudesh RathodNo ratings yet
- Early Development of Betta Splendens Under Stereomicroscopy and Scanning Electron MicrosDocument11 pagesEarly Development of Betta Splendens Under Stereomicroscopy and Scanning Electron Micros0 9No ratings yet
- 1962, Waddington, How Animals DevelopDocument148 pages1962, Waddington, How Animals DevelopLuis Ossa FuentesNo ratings yet
- Syllabus For B. Sc. Zoology (Honours & General) 2016: University of CalcuttaDocument25 pagesSyllabus For B. Sc. Zoology (Honours & General) 2016: University of CalcuttaRaajNo ratings yet
- Cleavage, Blastulation and Gastrulation Lab ReportDocument8 pagesCleavage, Blastulation and Gastrulation Lab ReportJOSHUA MOLO100% (1)
- BIOLOGY Form 5 Chapter 4Document65 pagesBIOLOGY Form 5 Chapter 4Shephard Png88% (8)
- Chick EmbryologyDocument55 pagesChick EmbryologySushma MishraNo ratings yet
- Fertilisasi, Implantasi and EmbriogenesisDocument57 pagesFertilisasi, Implantasi and EmbriogenesisjeinpratpongNo ratings yet
- Vertebrate Embryology - Randel PDFDocument399 pagesVertebrate Embryology - Randel PDFrandelNo ratings yet
- Four Year Zoology ProgrammeDocument16 pagesFour Year Zoology ProgrammeAmod KumarNo ratings yet
- Histology Lab GuideDocument96 pagesHistology Lab GuidevrindaNo ratings yet
- Development of AmphioxusDocument59 pagesDevelopment of AmphioxusDyah Ayu PuspitasariNo ratings yet
- Study of flower pollination and gamete developmentDocument6 pagesStudy of flower pollination and gamete developmentkavikiran60% (5)
- Development of Frog Umanga Chapagain Read OnlyDocument9 pagesDevelopment of Frog Umanga Chapagain Read OnlySubarna PudasainiNo ratings yet