You might also like
- Notes On EmbryologyDocument30 pagesNotes On EmbryologyArwa100% (9)
- Muscles of Head, Face & NeckDocument134 pagesMuscles of Head, Face & NeckDMNo ratings yet
- Orthopaedics and Trauma for Medical Students and Junior ResidentsFrom EverandOrthopaedics and Trauma for Medical Students and Junior ResidentsRating: 5 out of 5 stars5/5 (3)
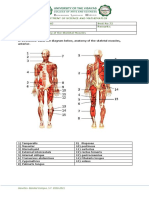
- Activity 3 - Human AnatomyDocument3 pagesActivity 3 - Human AnatomyNIRAJ PATELNo ratings yet
- แกะเทป Developmental of Musculoskeletal SystemDocument50 pagesแกะเทป Developmental of Musculoskeletal Systemjulesarojinee100% (1)
- PlacentaDocument36 pagesPlacentaAbdullah HussainyNo ratings yet
- Fetal and Placental Development - FETAL Sac, Umbilical Cord .Abnormalities of PlacentaDocument150 pagesFetal and Placental Development - FETAL Sac, Umbilical Cord .Abnormalities of PlacentaKripa SusanNo ratings yet
- Human Embryonic DevelopmentDocument8 pagesHuman Embryonic DevelopmentGargi BhattacharyaNo ratings yet
- Chapter 2 - General Characteristics of The SpineDocument50 pagesChapter 2 - General Characteristics of The SpineveronicalovirgenNo ratings yet
- Sclerotome Dermatome MyotomeDocument5 pagesSclerotome Dermatome Myotomeerick evanderNo ratings yet
- Embryology of Musculoskeletal SystemDocument14 pagesEmbryology of Musculoskeletal SystemdiahNo ratings yet
- EmbryologyDocument26 pagesEmbryologyMeer BabanNo ratings yet
- Cell Profile Report - Nisal Karawita - Nlkar1Document12 pagesCell Profile Report - Nisal Karawita - Nlkar1Lochana Karawita100% (1)
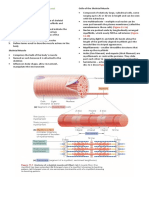
- Skeletal Muscle Cells and Their Packaging Into MusclesDocument7 pagesSkeletal Muscle Cells and Their Packaging Into MusclesDiana Rose SaludoNo ratings yet
- Fertilization and ImplantationDocument2 pagesFertilization and ImplantationAngelica AycardoNo ratings yet
- EmbryologyDocument68 pagesEmbryologyBj Larracas100% (1)
- Lecture 3: Musculoskeletal Development Slide 1Document9 pagesLecture 3: Musculoskeletal Development Slide 1Dionix CruzNo ratings yet
- Dev Muscular SystemDocument61 pagesDev Muscular Systemamandagnaw19No ratings yet
- Sistem Otot: Zuliyati RohmahDocument27 pagesSistem Otot: Zuliyati Rohmahnur90No ratings yet
- Development of Musculoskeletal SystemDocument25 pagesDevelopment of Musculoskeletal Systemwmdpr4x64fNo ratings yet
- Embryology Muscularskeletal SystemDocument32 pagesEmbryology Muscularskeletal SystemSurajul AudaNo ratings yet
- Skeletal Muscle Development and Regeneration: Concise ReviewDocument12 pagesSkeletal Muscle Development and Regeneration: Concise Reviewtesfaye mekonnenNo ratings yet
- Muscular System-11 (Muhadharaty)Document13 pagesMuscular System-11 (Muhadharaty)maysanitmaysanNo ratings yet
- Muscular SystemDocument18 pagesMuscular SystemAdan ImanNo ratings yet
- Development of Muscular System AtfDocument3 pagesDevelopment of Muscular System AtfKim Sa-buNo ratings yet
- Elective EntropyDocument21 pagesElective EntropyDavidroy IRITSNo ratings yet
- 26 - Development of Muscular SystemDocument12 pages26 - Development of Muscular SystemDr.B.B.GosaiNo ratings yet
- Musculo-Skeletal System: (Trunk, Limbs, and Head)Document52 pagesMusculo-Skeletal System: (Trunk, Limbs, and Head)kapwattNo ratings yet
- Caffey's Pediatric Diagnostic ImagingDocument8 pagesCaffey's Pediatric Diagnostic ImagingHafiz DjosanNo ratings yet
- Embryo Syst 1. Dev of Muscle and Cavities Mb2Document26 pagesEmbryo Syst 1. Dev of Muscle and Cavities Mb2Liz NGALEUNo ratings yet
- Myocytes Cell Profile ReportDocument11 pagesMyocytes Cell Profile ReportAreesha FatimaNo ratings yet
- Dev't of MuskuloskeletalDocument70 pagesDev't of MuskuloskeletalMilkoo sabaNo ratings yet
- Embryology and Anatomy: Spine/Spinal CordDocument5 pagesEmbryology and Anatomy: Spine/Spinal CordFernandoo CunhaNo ratings yet
- Dermatome: Embryo Tendons BoneDocument2 pagesDermatome: Embryo Tendons Bonetiko morchadzeNo ratings yet
- Fungsi Sistem OtotDocument8 pagesFungsi Sistem OtotMahesha RamadhaniNo ratings yet
- Lecture 13-Development of Skeletal & Muscular SystemDocument7 pagesLecture 13-Development of Skeletal & Muscular Systemtesfaye mekonnenNo ratings yet
- Development of The Musculoskeletal System: DR - Sahar HafeezDocument24 pagesDevelopment of The Musculoskeletal System: DR - Sahar HafeezDemuel Dee L. BertoNo ratings yet
- Axial Muscle Development in FishDocument7 pagesAxial Muscle Development in FishMoisés Aldana AcostaNo ratings yet
- MusclesssDocument4 pagesMusclesssLiam Jacque LapuzNo ratings yet
- Muscle Tissue SummaryDocument2 pagesMuscle Tissue SummaryLindenScholesNo ratings yet
- Mesodermal Tissue and Its Derivatives: MyocoelDocument62 pagesMesodermal Tissue and Its Derivatives: Myocoelytterbium101No ratings yet
- Mesoderm Formation / Segmentation: Reading Assignment: SummaryDocument10 pagesMesoderm Formation / Segmentation: Reading Assignment: Summaryaghsal novanNo ratings yet
- Lecture 1 NotesDocument1 pageLecture 1 Notesgatorssoccer1313No ratings yet
- Kardong 154-170,462-464 Color FiguresDocument7 pagesKardong 154-170,462-464 Color FiguresNishat_Fatima_3779No ratings yet
- Weeks 3 - 8Document32 pagesWeeks 3 - 8Stefan HutsonNo ratings yet
- Embryonic Period 4-8weekDocument25 pagesEmbryonic Period 4-8weekaimi BatrisyiaNo ratings yet
- Submodule 2.5 - 2.6Document14 pagesSubmodule 2.5 - 2.6Zeejiah NelleNo ratings yet
- 1707721775Document19 pages1707721775atrayeeganguly8f39No ratings yet
- Locomotion and Movement VENDocument12 pagesLocomotion and Movement VENSREE GANESHNo ratings yet
- Gastrulation and Segmentation ofDocument20 pagesGastrulation and Segmentation ofAina AdesolaNo ratings yet
- The Neck: Overview and Objectives of This DissectionDocument5 pagesThe Neck: Overview and Objectives of This Dissectionapi-61200414No ratings yet
- 09 - Musculoskeletal DevelopmentDocument8 pages09 - Musculoskeletal DevelopmentAXA OFFICIALNo ratings yet
- K. Pengantar Muskulofisiologi 1, Dr. M. Djauhari W., AIF, PFK, Kamis 28 Oktober 2021Document116 pagesK. Pengantar Muskulofisiologi 1, Dr. M. Djauhari W., AIF, PFK, Kamis 28 Oktober 2021punthadewaNo ratings yet
- The Muscular TissueDocument23 pagesThe Muscular TissueMuhammad Salisu IliyasuNo ratings yet
- IB Singh Human Embryology Part 2Document547 pagesIB Singh Human Embryology Part 2Rajeshwari RajiNo ratings yet
- Anatomy, Head and Neck Frontalis Muscle ArticleDocument7 pagesAnatomy, Head and Neck Frontalis Muscle ArticleWindsurfingFinnNo ratings yet
- Musculature - Module 2Document37 pagesMusculature - Module 2CaityNo ratings yet
- Locomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)Document13 pagesLocomotion and Movement Class 11 Notes CBSE Biology Chapter 20 (PDF)ASHISH MALAVNo ratings yet
- Anatomy of Musculoskeletal and Integumentary Systems By:Mihretu JDocument53 pagesAnatomy of Musculoskeletal and Integumentary Systems By:Mihretu JAmanuel MaruNo ratings yet
- Elevage Des Veaux D'elevageDocument6 pagesElevage Des Veaux D'elevageabdelaziz CHELIGHOUMNo ratings yet
- Module 5. Muscle Tissue17Document7 pagesModule 5. Muscle Tissue17Mark Jay PagaduanNo ratings yet
- Generation of HeatDocument2 pagesGeneration of HeatFani Try Utami BoedaxRed'waterzzNo ratings yet
- Appendicular & LimbDocument29 pagesAppendicular & LimbPeter PrestonNo ratings yet
- NCLEX Review NotesDocument12 pagesNCLEX Review NotesWillington Cuaresma100% (1)
- Chordate E&D PDFDocument14 pagesChordate E&D PDFPatience HeduvorNo ratings yet
- Book - The Frog Its Reproduction and Development 17 - EmbryologyDocument12 pagesBook - The Frog Its Reproduction and Development 17 - EmbryologyJudy Anne Vitales GalizaNo ratings yet
- Fate Mapping - Gastrulation - Lec Notes - KMZRDocument27 pagesFate Mapping - Gastrulation - Lec Notes - KMZRSaad SakalNo ratings yet
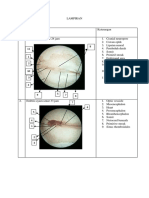
- LAMPIRAN Embrio AyamDocument3 pagesLAMPIRAN Embrio AyamdidinNo ratings yet
- Republic of The PhilippinesDocument5 pagesRepublic of The Philippineshatdog adadaNo ratings yet
- Human Reproduction 2 Notes (Class 12)Document6 pagesHuman Reproduction 2 Notes (Class 12)Ashok KumarNo ratings yet
- SYBSc Sem 4 Practical No. 10 Types of CleavageDocument4 pagesSYBSc Sem 4 Practical No. 10 Types of CleavageAnkit NariyaNo ratings yet
- Vet Obst Lecture 1 Pregnancy in Domestic AnimalsDocument53 pagesVet Obst Lecture 1 Pregnancy in Domestic AnimalsgnpobsNo ratings yet
- 4 - Embryology Lecture 4 (Week 2 of Development A)Document31 pages4 - Embryology Lecture 4 (Week 2 of Development A)Mostafa MostafaNo ratings yet
- Forms of Embryonic Primordia: Prof. Anthony Obioma NwaoparaDocument32 pagesForms of Embryonic Primordia: Prof. Anthony Obioma NwaoparamatthewNo ratings yet
- Correlation Seminar On Reproductive and Endocrine SystemDocument20 pagesCorrelation Seminar On Reproductive and Endocrine Systemnischal neupaneNo ratings yet
- The Developing Human Clinically Oriented Embryology 9th Edition Moore Persaud Torchia Test BankDocument24 pagesThe Developing Human Clinically Oriented Embryology 9th Edition Moore Persaud Torchia Test BankAmberLongpmtg100% (33)
- Development StagesDocument4 pagesDevelopment StagesSerban RalucaNo ratings yet
- Embriologia Del CerdoDocument11 pagesEmbriologia Del CerdoTadase Hotori0% (1)
- Embryonic Period 4-8weekDocument25 pagesEmbryonic Period 4-8weekaimi BatrisyiaNo ratings yet
- Development of The Nervous System 3rd Edition Sanes Test BankDocument35 pagesDevelopment of The Nervous System 3rd Edition Sanes Test Bankbateddivisor51zwed100% (20)
- Bio 109 Module 4Document18 pagesBio 109 Module 4Marites Maylanon0% (1)
- Pattern Formation in DrosophilaDocument17 pagesPattern Formation in DrosophilaAdwika DeoNo ratings yet
- Gas Tru LationDocument8 pagesGas Tru LationemmaqwashNo ratings yet
- Developmental Biology 10 Placental Structure and ClassificationDocument6 pagesDevelopmental Biology 10 Placental Structure and ClassificationPijush PramanikNo ratings yet
- Stages of Growth and Development: Before BirthDocument3 pagesStages of Growth and Development: Before BirthZowi Sian MedesNo ratings yet
- Data Pengamatan Embrio AyamDocument5 pagesData Pengamatan Embrio AyamdindaNo ratings yet
- Gastrulation (20-10-2019)Document16 pagesGastrulation (20-10-2019)kashif manzoorNo ratings yet
- OB M01 Ch1.1 Normal First TrimesterDocument29 pagesOB M01 Ch1.1 Normal First TrimesterDeshi SportsNo ratings yet
- Prenatal DevelopmentDocument3 pagesPrenatal DevelopmentJea Sheinelle G. DiazNo ratings yet