You might also like
- Co-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseFrom EverandCo-signal Molecules in T Cell Activation: Immune Regulation in Health and DiseaseMiyuki AzumaNo ratings yet
- Lymphocyte Activation 25 Dan 27 Sep 2017Document39 pagesLymphocyte Activation 25 Dan 27 Sep 2017Kohita R PerdanaNo ratings yet
- CH 7 Antibody 7E03Document1 pageCH 7 Antibody 7E03JUSASBNo ratings yet
- Introduction To Immunology and Immunotherapy: Section 1Document13 pagesIntroduction To Immunology and Immunotherapy: Section 1ZairullahMighfazaNo ratings yet
- Session 16-2 B Cell Response (T Independent)Document4 pagesSession 16-2 B Cell Response (T Independent)Satwik korukondaNo ratings yet
- Cooperari Celulare Si Mecanisme in Raspunsul Imun Celular Si UmoralDocument12 pagesCooperari Celulare Si Mecanisme in Raspunsul Imun Celular Si UmoralMariam DalatyNo ratings yet
- Adaptive Immunity: Wesam Mohammed EmharibDocument15 pagesAdaptive Immunity: Wesam Mohammed Emharibنسيم حامدNo ratings yet
- B Cell Activation and the Mechanisms of Humoral ImmunityDocument9 pagesB Cell Activation and the Mechanisms of Humoral ImmunityPavel NesmiyanovNo ratings yet
- Ctivation of Ells: Abul Abbas, MD EmailDocument4 pagesCtivation of Ells: Abul Abbas, MD EmailDr.Santosh KumarNo ratings yet
- C-12-Tcell and Bcell ActivationDocument43 pagesC-12-Tcell and Bcell Activationlinnet17No ratings yet
- Cooperari CelulareDocument12 pagesCooperari Celularemiruna_cNo ratings yet
- Humoral Immunity B CellDocument11 pagesHumoral Immunity B CellAbod AbodNo ratings yet
- B Cell ActivationDocument62 pagesB Cell ActivationAYSHAMOHD88No ratings yet
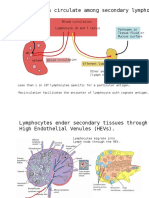
- Mature T Cells Circulate Among Secondary Lymphoid Tissues.: Blood CirculationDocument30 pagesMature T Cells Circulate Among Secondary Lymphoid Tissues.: Blood CirculationNovi LatifaNo ratings yet
- Immunology Mid-ExamDocument11 pagesImmunology Mid-ExamNgMinhHaiNo ratings yet
- Adaptive Immunity IDocument29 pagesAdaptive Immunity Ixfxw5m5bczNo ratings yet
- Adaptive Immune ResponseDocument13 pagesAdaptive Immune ResponseMuhammad FaheemNo ratings yet
- Immunology Study Packet #1 AnswersDocument6 pagesImmunology Study Packet #1 AnswersAnnTranNo ratings yet
- Fundamentals of Pathology The Adaptive Immune Response T CellsDocument41 pagesFundamentals of Pathology The Adaptive Immune Response T CellsOsman MNo ratings yet
- Lecture 3Document5 pagesLecture 3zeynep sevinçNo ratings yet
- Antigen Presenting Cells: Parbondeep SaikiaDocument20 pagesAntigen Presenting Cells: Parbondeep SaikiaParbondeep SaikiaNo ratings yet
- The Role of Complement Proteins in B-Cell Activation C3d ("Second Signals") 1. Antigen Receptor-Mediated Signal Transduction in B-LymphocytesDocument5 pagesThe Role of Complement Proteins in B-Cell Activation C3d ("Second Signals") 1. Antigen Receptor-Mediated Signal Transduction in B-LymphocytesJaz MnNo ratings yet
- Linfocitos B y T-DepuratedDocument18 pagesLinfocitos B y T-DepuratedByron Emerson Gonzales GonzalesNo ratings yet
- Chapter 16 Homework Part IIDocument6 pagesChapter 16 Homework Part IIKvn4N6No ratings yet
- 07cell Cell InteractionsDocument9 pages07cell Cell InteractionsAhmed Goma'aNo ratings yet
- Immune SystemDocument32 pagesImmune Systemanupama menonNo ratings yet
- Immunology Review: Key ConceptsDocument6 pagesImmunology Review: Key ConceptsFatmala Umi MaisarahNo ratings yet
- Immunology in Haematology (Part 1)Document48 pagesImmunology in Haematology (Part 1)kiedd_04100% (4)
- Lecture - Cell Mediated ImmunityDocument34 pagesLecture - Cell Mediated Immunityrockyrawat01012003No ratings yet
- 9.MHC in Immune Response-2Document16 pages9.MHC in Immune Response-2alhyarylyn3No ratings yet
- Full Download Basic Immunology Functions and Disorders of The Immune System 4th Edition Abbas Test BankDocument36 pagesFull Download Basic Immunology Functions and Disorders of The Immune System 4th Edition Abbas Test Bankpasakazinum100% (34)
- Chapter 9: The Humoral Immune Response I. B Cell Activation and Antibody ProductionDocument12 pagesChapter 9: The Humoral Immune Response I. B Cell Activation and Antibody ProductionPaul DavisNo ratings yet
- Reference: pg.24: Clinical Immunology & Serology: A Laboratory Perspective Christine Dorresteyn StevensDocument1 pageReference: pg.24: Clinical Immunology & Serology: A Laboratory Perspective Christine Dorresteyn StevensLorelyn Lacusong CendañaNo ratings yet
- Tolerance 1Document7 pagesTolerance 1Asmaa RayanNo ratings yet
- 44 pdl1Document3 pages44 pdl1bawoji1763No ratings yet
- Immuno First - Past Years QsDocument48 pagesImmuno First - Past Years QsAbdallah Essam Al-Zireeni67% (3)
- Antigens, Hapteins, Immunogens Lectures 10.1.06Document32 pagesAntigens, Hapteins, Immunogens Lectures 10.1.06CLEMENTNo ratings yet
- Fundamentals of Immunology - Death by Friendly FireDocument145 pagesFundamentals of Immunology - Death by Friendly FireAngel FeresiaNo ratings yet
- A Public BCR Present in A Unique Dual-Receptor-Expressing Lymphocyte From Type 1 Diabetes Patients Encodes A Potent T Cell AutoantigenDocument34 pagesA Public BCR Present in A Unique Dual-Receptor-Expressing Lymphocyte From Type 1 Diabetes Patients Encodes A Potent T Cell AutoantigenAmrendra SinghNo ratings yet
- Sanchez Trincado2017Document15 pagesSanchez Trincado2017Andres JarrinNo ratings yet
- Cell-Mediated Immunity: A. Antigen - Specific CellsDocument7 pagesCell-Mediated Immunity: A. Antigen - Specific CellsandindlNo ratings yet
- Week 1 - Jan 27 Lesson Plan: Innate Immunity Adaptive ImmunityDocument40 pagesWeek 1 - Jan 27 Lesson Plan: Innate Immunity Adaptive ImmunitychocobuniNo ratings yet
- Inmunología de las vacunas: Respuestas a la vacuna de células TDocument4 pagesInmunología de las vacunas: Respuestas a la vacuna de células TMontserrat GarciaNo ratings yet
- B Cells T Cells IgsDocument84 pagesB Cells T Cells IgsKhatrinaNo ratings yet
- T Cells & B Cells: 6.1 - Antigen Processing and PresentationDocument6 pagesT Cells & B Cells: 6.1 - Antigen Processing and PresentationAmanda AmiraultNo ratings yet
- Immunopathology: Normal Immune ResponseDocument13 pagesImmunopathology: Normal Immune ResponseChinedu H. DuruNo ratings yet
- Immunology MSQDocument2 pagesImmunology MSQNaing Lin SoeNo ratings yet
- 4801 - Activation of T CellDocument28 pages4801 - Activation of T CellaamohonbdNo ratings yet
- Chapter Seven: Immunity Response Induction: Granulocyte ExocytosisDocument4 pagesChapter Seven: Immunity Response Induction: Granulocyte ExocytosisNur LiyanaNo ratings yet
- T Lymphocyte Activation: Key Steps and RegulationDocument15 pagesT Lymphocyte Activation: Key Steps and Regulationnadeem banyhanyNo ratings yet
- CMI- Cell Mediated Immunity OverviewDocument34 pagesCMI- Cell Mediated Immunity Overviewlion2chNo ratings yet
- Cell Mediated Immunity: Department of Parasitology, Medical Faculty, Hasanuddin UniversityDocument64 pagesCell Mediated Immunity: Department of Parasitology, Medical Faculty, Hasanuddin Universityintan yusufNo ratings yet
- Immunology in Haematology (Part 2)Document55 pagesImmunology in Haematology (Part 2)kiedd_04100% (4)
- Immunologia zakažeä-ENG (Automatycznie Zapisany) - KopiaDocument41 pagesImmunologia zakažeä-ENG (Automatycznie Zapisany) - KopiaAssoNo ratings yet
- Immunology 2Document30 pagesImmunology 2ukashazam19No ratings yet
- 5 adaptive immunity (1)Document35 pages5 adaptive immunity (1)ozerbilge24No ratings yet
- Genetic Basis of ImmunodeficiencyDzDocument2 pagesGenetic Basis of ImmunodeficiencyDzcoolradishNo ratings yet
- Immune Regulation and ToleranceDocument33 pagesImmune Regulation and ToleranceAniruddha RoyNo ratings yet
- Section 13 B Cell Mediated Humoral ImmunityDocument33 pagesSection 13 B Cell Mediated Humoral ImmunityManaswiniNo ratings yet
- Humoral Immune System-MelisaDocument55 pagesHumoral Immune System-Melisameydiana ivankaNo ratings yet
- Investigating Respiratory Illness in ChildrenDocument19 pagesInvestigating Respiratory Illness in ChildrenOxana TurcuNo ratings yet
- Scope Guideline Pain in ChildrenDocument11 pagesScope Guideline Pain in ChildrenKomang Prawira Nata NugrahaNo ratings yet
- Pain Management in Infants - Children - AdolescentsDocument9 pagesPain Management in Infants - Children - AdolescentsOxana Turcu100% (1)
- Allergic Bronchopulmonary Aspergillosis Clinical Care Guidelines - CF FoundationDocument8 pagesAllergic Bronchopulmonary Aspergillosis Clinical Care Guidelines - CF FoundationOxana TurcuNo ratings yet
- Paediatric AsthmaDocument246 pagesPaediatric AsthmaOxana TurcuNo ratings yet
- Convulsion Febril - Febrile Seizures Risks, Evaluation, and Prognosis - Am Fam Physician. 2012Document5 pagesConvulsion Febril - Febrile Seizures Risks, Evaluation, and Prognosis - Am Fam Physician. 2012Nataly Zuluaga VilladaNo ratings yet
- Acute Exacerbations of Pulmonary DiseasesDocument258 pagesAcute Exacerbations of Pulmonary DiseasesOxana TurcuNo ratings yet
- ERS Statement On Protracted Bacterial BronchitisDocument18 pagesERS Statement On Protracted Bacterial BronchitisOxana TurcuNo ratings yet
- Shaken Baby Syndrome PDFDocument12 pagesShaken Baby Syndrome PDFOxana TurcuNo ratings yet
- Down Syndrome Cognitive DisabilityDocument2 pagesDown Syndrome Cognitive DisabilityOxana TurcuNo ratings yet
- RthwertDocument17 pagesRthwertAnnisa FujiantiNo ratings yet
- Borg Scale in Children Older Than 9 With CFDocument5 pagesBorg Scale in Children Older Than 9 With CFOxana TurcuNo ratings yet
- Rath BarbaraDocument17 pagesRath Barbarasmansa123No ratings yet
- DBD Fever and RashDocument11 pagesDBD Fever and RashSa 'ng WijayaNo ratings yet
- Dalhousie Dyspnea ScalesDocument7 pagesDalhousie Dyspnea ScalesOxana TurcuNo ratings yet
- Enfermedad Febril en Niños Menores de 5 AñosDocument43 pagesEnfermedad Febril en Niños Menores de 5 AñosMarco RiveraNo ratings yet
- MCN Outpatient Care of Children With Acute Malnutrition Trainers ManualDocument69 pagesMCN Outpatient Care of Children With Acute Malnutrition Trainers ManualOxana TurcuNo ratings yet
- Breastfeeding - The Essential PrinciplesDocument11 pagesBreastfeeding - The Essential PrinciplesOxana TurcuNo ratings yet
- Antibiotic Prescribing in 2016Document9 pagesAntibiotic Prescribing in 2016Oxana TurcuNo ratings yet
- InTech-Arrhythmias in Children and Young AdultsDocument36 pagesInTech-Arrhythmias in Children and Young Adultsmadalina8No ratings yet
- Radiological Features of BronchiectasisDocument24 pagesRadiological Features of BronchiectasisOxana TurcuNo ratings yet
- EngDocument256 pagesEngClaudia BocanegraNo ratings yet
- Pediatric Respiratory EmergenciesDocument68 pagesPediatric Respiratory EmergenciesPAKINUGNo ratings yet
- Host-Microbe Interactions in Distal AirwaysDocument14 pagesHost-Microbe Interactions in Distal AirwaysOxana TurcuNo ratings yet
- Non CF Bronchiectasis GuidelineDocument64 pagesNon CF Bronchiectasis GuidelineJavier Huanca BaezNo ratings yet
- Anitbiotic Treatment in Cystic FibrosisDocument76 pagesAnitbiotic Treatment in Cystic FibrosisOxana TurcuNo ratings yet
- Radiological Features of BronchiectasisDocument24 pagesRadiological Features of BronchiectasisOxana TurcuNo ratings yet
- InTech-Arrhythmias in Children and Young AdultsDocument36 pagesInTech-Arrhythmias in Children and Young Adultsmadalina8No ratings yet
- Primary ImmunodefDocument12 pagesPrimary ImmunodefOxana TurcuNo ratings yet
- Akeela House Intake Packet RDocument11 pagesAkeela House Intake Packet RDamonNo ratings yet
- Effectiveness of T-Piece Resuscitator Versus Self-Inflating Bag During Birth Resuscitation in Very Low Birth Weight InfantsDocument7 pagesEffectiveness of T-Piece Resuscitator Versus Self-Inflating Bag During Birth Resuscitation in Very Low Birth Weight InfantsRaja SekarNo ratings yet
- January 2022 BehavioursDocument8 pagesJanuary 2022 Behavioursdhopat kalpeshNo ratings yet
- FomaDocument1,218 pagesFomaosamaeNo ratings yet
- Addicted To Phones - Reading AssignmentDocument4 pagesAddicted To Phones - Reading AssignmentLilith PersephoneNo ratings yet
- Timeless Beauty - Over 100 Tips, Secrets, and Shortcuts To Looking Great (PDFDrive)Document488 pagesTimeless Beauty - Over 100 Tips, Secrets, and Shortcuts To Looking Great (PDFDrive)Isabel Matta100% (1)
- Instability of The Distal Radioulnar JointDocument12 pagesInstability of The Distal Radioulnar JointOryza SatriaNo ratings yet
- SV The Coronation of King Charles IIIDocument9 pagesSV The Coronation of King Charles IIIMaria GromovaNo ratings yet
- VA - DQA Specialist IBTCIDocument5 pagesVA - DQA Specialist IBTCIT. Woibah SuwoNo ratings yet
- AugerTorque European Trencher Operators ManualDocument23 pagesAugerTorque European Trencher Operators ManualПавел ХромовNo ratings yet
- Expected Coding and Decoding Questions For Ibps Po MainsDocument11 pagesExpected Coding and Decoding Questions For Ibps Po Mainsvirat kohli86% (7)
- Students Personal Data Sheet FormDocument1 pageStudents Personal Data Sheet FormL 1No ratings yet
- Medical Certificate: Healthservices@lnu - Edu.phDocument3 pagesMedical Certificate: Healthservices@lnu - Edu.phNiko ChavezNo ratings yet
- College of Liberal Arts University of LuzonDocument9 pagesCollege of Liberal Arts University of LuzonChristianne Joyse MerreraNo ratings yet
- Professional Teachers (Secondary) 10-2022 FilipinoDocument26 pagesProfessional Teachers (Secondary) 10-2022 FilipinoPRC Baguio100% (1)
- The Relationship Between Lower Intelligence Crime and Custodial Outcomes A Brief Literary Review of A Vulnerable GroupDocument16 pagesThe Relationship Between Lower Intelligence Crime and Custodial Outcomes A Brief Literary Review of A Vulnerable GroupSteveNo ratings yet
- New Pattern QP-1Document50 pagesNew Pattern QP-1Unknown cool KidNo ratings yet
- Lecture 3.2.3: Erection III: Steel Construction: Fabrication and ErectionDocument4 pagesLecture 3.2.3: Erection III: Steel Construction: Fabrication and ErectionMihajloDjurdjevicNo ratings yet
- Chapter 1Document2 pagesChapter 1longaitiNo ratings yet
- Government Issues Relief Guidelines for Victims of AtrocitiesDocument4 pagesGovernment Issues Relief Guidelines for Victims of AtrocitiesDept . WD&CW Andhra PradeshNo ratings yet
- RA-032739 - MASTER PLUMBER - Lucena - 2-2023Document7 pagesRA-032739 - MASTER PLUMBER - Lucena - 2-2023Bon Opeña GorobaoNo ratings yet
- Re-Opening Plan For The 2020-2021 School YearDocument18 pagesRe-Opening Plan For The 2020-2021 School YearNews 8 WROCNo ratings yet
- End of Life Care GuideDocument79 pagesEnd of Life Care GuideAbcd TolibasNo ratings yet
- Hope Week 2Document5 pagesHope Week 2Bea CabacunganNo ratings yet
- Artikel Bahasa Inggris Dengan Topik BerbedaDocument2 pagesArtikel Bahasa Inggris Dengan Topik Berbedaputra haqiqiNo ratings yet
- HSE and SOP ON EMBARKATIONDocument5 pagesHSE and SOP ON EMBARKATIONNajmirzan IsraNo ratings yet
- Healthlittoolkit2 Tool2aDocument7 pagesHealthlittoolkit2 Tool2aRenzz Irven PlotadoNo ratings yet
- SPTL Ohs&e 40 Ohse MPR AbrarDocument22 pagesSPTL Ohs&e 40 Ohse MPR AbrarRanjeet Kumar SinghNo ratings yet
- Prasnottara English For AmazonDocument127 pagesPrasnottara English For AmazonAnand PendyalaNo ratings yet
- Modals of Suggestion HomeworkDocument2 pagesModals of Suggestion HomeworkmateoNo ratings yet