You might also like
- Organ Anatomy Notes 1Document8 pagesOrgan Anatomy Notes 1ELIANA CARIS CABILDONo ratings yet
- Merchant Accounts Are Bank Accounts That Allow Your Business To Accept Card Payments From CustomersDocument43 pagesMerchant Accounts Are Bank Accounts That Allow Your Business To Accept Card Payments From CustomersRohit Kumar Baghel100% (1)
- 4th Conference ParticipantsDocument14 pages4th Conference ParticipantsmaxNo ratings yet
- Special Senses Physiology of Hearing and Balance: Dr. CalaquianDocument6 pagesSpecial Senses Physiology of Hearing and Balance: Dr. CalaquianHanako Sasaki AranillaNo ratings yet
- Pa Ears-Nose-MouthDocument108 pagesPa Ears-Nose-MouthDenniellePaulineKwonNo ratings yet
- Anatomy of EarDocument42 pagesAnatomy of EarHesti hasanNo ratings yet
- Patho Chaopter 2Document21 pagesPatho Chaopter 2KC White Dela RosaNo ratings yet
- VISCOLAM202 D20 Acrylic 20 Thickeners 202017Document33 pagesVISCOLAM202 D20 Acrylic 20 Thickeners 202017Oswaldo Manuel Ramirez MarinNo ratings yet
- MS2 EarsDocument8 pagesMS2 Earswieka mawieNo ratings yet
- HSB - Head: Anterior: Supraorbital Margin Posterior: Superior Nuchal LineDocument19 pagesHSB - Head: Anterior: Supraorbital Margin Posterior: Superior Nuchal LineRaki IliganNo ratings yet
- DWDMDocument41 pagesDWDMKarthik KompelliNo ratings yet
- The Ear. The Helix. The External Auditory Canal and Tympanic Membrane. Air and Bone JoiningDocument40 pagesThe Ear. The Helix. The External Auditory Canal and Tympanic Membrane. Air and Bone JoiningHarshit BhardwajNo ratings yet
- Anatomy of The EarDocument41 pagesAnatomy of The EarDenny Briant100% (1)
- Antonov 225 - The Largest - Airliner in The WorldDocument63 pagesAntonov 225 - The Largest - Airliner in The WorldFridayFunStuffNo ratings yet
- Physiology of Hearing OtolaryngologyDocument26 pagesPhysiology of Hearing OtolaryngologyDr.Sherif Bugnah88% (8)
- 3.1 One Pager The Ear - MemoDocument1 page3.1 One Pager The Ear - MemoRudzi UdziNo ratings yet
- Lesson 1 The Human Ear Structure and Function LessonDocument7 pagesLesson 1 The Human Ear Structure and Function LessonCarvia CusNo ratings yet
- Anatomy + Objective PhenomenaDocument31 pagesAnatomy + Objective PhenomenaHoussemNo ratings yet
- Module 4 PPT The Ear 2022 EditDocument43 pagesModule 4 PPT The Ear 2022 Editaesthete541No ratings yet
- Xibraho: AuditoryDocument6 pagesXibraho: AuditoryYousuf skNo ratings yet
- Mechanism of Hearing PDFDocument6 pagesMechanism of Hearing PDFShruthi DanielNo ratings yet
- Assessing EarsDocument11 pagesAssessing EarsRhea Mae Visda RoaNo ratings yet
- 05 - Hearing and BalanceDocument23 pages05 - Hearing and BalancearonsnorrasonNo ratings yet
- Special Senses Part II - PearsonsDocument5 pagesSpecial Senses Part II - Pearsonsblando.shaneNo ratings yet
- Signal Processing For Auditory SystemsDocument108 pagesSignal Processing For Auditory SystemsKarthikeya SharmaNo ratings yet
- Physiology of Auditory SystemDocument26 pagesPhysiology of Auditory SystemVictor EnachiNo ratings yet
- Human EarDocument21 pagesHuman EarDivya RajendranNo ratings yet
- Appendicular Skeleton 1-1Document117 pagesAppendicular Skeleton 1-1Anania EmmanuelNo ratings yet
- Physiology of EarDocument80 pagesPhysiology of EarPhysiology by Dr Raghuveer50% (2)
- Ossicles: The Malleus, The Incus, and The StapesDocument6 pagesOssicles: The Malleus, The Incus, and The StapesLili M.No ratings yet
- Vertebral Column and Contents of The Vertebral CanalDocument6 pagesVertebral Column and Contents of The Vertebral CanalCay KaiNo ratings yet
- KC - Longitudinal and Transverse - KnowDocument50 pagesKC - Longitudinal and Transverse - KnowT WNo ratings yet
- 4bi 1le Active RecallDocument4 pages4bi 1le Active RecallAlexandryaHaleNo ratings yet
- Q1 W8 M3 MusicDocument4 pagesQ1 W8 M3 MusicChristian Don VelasquezNo ratings yet
- Sense OrgansDocument3 pagesSense OrgansAabhaNo ratings yet
- Knowledge Organiser: Properties of Waves Sound Waves HearingDocument1 pageKnowledge Organiser: Properties of Waves Sound Waves HearingLuisGamesNo ratings yet
- The EarDocument103 pagesThe Eardharmendra kirarNo ratings yet
- C1 Atlas C2 Axis C3 - C6 Typical Cervical Vertebra C7 Vertebra ProminensDocument4 pagesC1 Atlas C2 Axis C3 - C6 Typical Cervical Vertebra C7 Vertebra ProminensmiallyannaNo ratings yet
- Physiology of The Auditory SystemDocument47 pagesPhysiology of The Auditory Systemmkct111No ratings yet
- The Sense of HearingDocument11 pagesThe Sense of HearingAfdalia AmandaNo ratings yet
- The Ear Study GuideDocument2 pagesThe Ear Study GuideBrianne EarnestNo ratings yet
- Guyton and Hall Medical Physiology Copy 2Document19 pagesGuyton and Hall Medical Physiology Copy 2Ful VianaNo ratings yet
- CNS 4 GoodDocument16 pagesCNS 4 GoodJeff ParkNo ratings yet
- Hearing Physiology - GuytonDocument11 pagesHearing Physiology - GuytonDuc HaNo ratings yet
- Hearing Outline: - AnatomyDocument82 pagesHearing Outline: - AnatomyFadil BachmidNo ratings yet
- Organ Anatomy Notes 2Document11 pagesOrgan Anatomy Notes 2ELIANA CARIS CABILDONo ratings yet
- Finals EarsDocument9 pagesFinals EarsMonica JubaneNo ratings yet
- GA&E 5 - Intro To Arthrology & Osteology IIIDocument51 pagesGA&E 5 - Intro To Arthrology & Osteology IIISu ZikaiNo ratings yet
- Vertebral Column and ContentsDocument5 pagesVertebral Column and ContentsJewelNo ratings yet
- Human Senses and Human System : Auditory SenseDocument9 pagesHuman Senses and Human System : Auditory SenseJolly Ann PerdonNo ratings yet
- Management of VertigoDocument86 pagesManagement of VertigowilaNo ratings yet
- Facial BonesDocument146 pagesFacial BonesPavan100% (1)
- 5 Assessing EarsDocument18 pages5 Assessing EarsAnecito BrillantesNo ratings yet
- Shoulder Problem Evaluation: 6 Articulations or JointsDocument4 pagesShoulder Problem Evaluation: 6 Articulations or JointsMARY GIZETH GACHONo ratings yet
- Hearing (Inner Ear)Document15 pagesHearing (Inner Ear)ShaheerNo ratings yet
- Nose and Paranasal SinusesDocument4 pagesNose and Paranasal SinusesAlloiBialbaNo ratings yet
- SAS - Session 7 - PSY079 - Intro To PsychologyDocument8 pagesSAS - Session 7 - PSY079 - Intro To PsychologyLordelee Mae N PamaylaonNo ratings yet
- Anatomy and Physiology of Nose and PNSDocument6 pagesAnatomy and Physiology of Nose and PNSChalsey Jene LorestoNo ratings yet
- Voice 1Document131 pagesVoice 1Sumanth TjNo ratings yet
- LO SSS Week 5Document8 pagesLO SSS Week 5Tony StarkNo ratings yet
- VB Anatomy of Hearing 061119Document13 pagesVB Anatomy of Hearing 061119Marcos Aurélio RodriguesNo ratings yet
- Grade 12 Life Science The Ear PDFDocument5 pagesGrade 12 Life Science The Ear PDFLethabo Mngomezulu100% (1)
- The Stimulus Input: Sound WavesDocument23 pagesThe Stimulus Input: Sound Wavesapi-300762638No ratings yet
- Trans Biochem Chapter 36 - 394-399Document4 pagesTrans Biochem Chapter 36 - 394-399KC White Dela RosaNo ratings yet
- Light ReflexDocument2 pagesLight ReflexKC White Dela RosaNo ratings yet
- Gas DiffusionDocument1 pageGas DiffusionKC White Dela RosaNo ratings yet
- Hippocratic Oath: One of The Oldest Binding Documents in History, The OathDocument4 pagesHippocratic Oath: One of The Oldest Binding Documents in History, The OathKC White Dela RosaNo ratings yet
- Exercise BPDocument1 pageExercise BPKC White Dela RosaNo ratings yet
- Trans Neuro 141-145Document4 pagesTrans Neuro 141-145KC White Dela RosaNo ratings yet
- Social FactorsDocument11 pagesSocial FactorsKC White Dela RosaNo ratings yet
- Function of MicrocirculationDocument9 pagesFunction of MicrocirculationKC White Dela RosaNo ratings yet
- Gas DiffusionDocument1 pageGas DiffusionKC White Dela RosaNo ratings yet
- Gerald Albert GercayoDocument1 pageGerald Albert GercayoKC White Dela RosaNo ratings yet
- Diagnosis and Treatment OptionsDocument1 pageDiagnosis and Treatment OptionsKC White Dela RosaNo ratings yet
- Inflammation and RepairDocument21 pagesInflammation and RepairKC White Dela RosaNo ratings yet
- Exercise BPDocument1 pageExercise BPKC White Dela RosaNo ratings yet
- Location of Nonpolar Amino Acids in ProteinsDocument1 pageLocation of Nonpolar Amino Acids in ProteinsKC White Dela RosaNo ratings yet
- The Requirement For PPEDocument1 pageThe Requirement For PPEKC White Dela RosaNo ratings yet
- Thyroid GlandDocument2 pagesThyroid GlandKC White Dela RosaNo ratings yet
- Amino Acid HighlightsDocument2 pagesAmino Acid HighlightsKC White Dela RosaNo ratings yet
- General Principles of Gastrointestinal Motility: Alimentary TractDocument6 pagesGeneral Principles of Gastrointestinal Motility: Alimentary TractKC White Dela Rosa100% (1)
- What Is HepatitisDocument7 pagesWhat Is HepatitisKC White Dela RosaNo ratings yet
- Location of Nonpolar Amino Acids in ProteinsDocument1 pageLocation of Nonpolar Amino Acids in ProteinsKC White Dela RosaNo ratings yet
- FMP-Finals Sec A&B: Email AddressDocument15 pagesFMP-Finals Sec A&B: Email AddressKC White Dela RosaNo ratings yet
- Allosteric ModificationDocument1 pageAllosteric ModificationKC White Dela RosaNo ratings yet
- TRIGEMINALDocument3 pagesTRIGEMINALKC White Dela RosaNo ratings yet
- ANATOMY First Sem Final ExamDocument25 pagesANATOMY First Sem Final ExamKC White Dela RosaNo ratings yet
- Action PotentialDocument1 pageAction PotentialKC White Dela RosaNo ratings yet
- Introduction To Endocrinology (Chapter 75) : Lacaden, LMGCDocument4 pagesIntroduction To Endocrinology (Chapter 75) : Lacaden, LMGCKC White Dela Rosa100% (1)
- Physio LAB Prelim Exam 2nd Sem EditedDocument10 pagesPhysio LAB Prelim Exam 2nd Sem EditedKC White Dela RosaNo ratings yet
- Yung Mga Na Ka Red Na Questions, Wrong Ba Yung Nakablack Na Answer?????Document44 pagesYung Mga Na Ka Red Na Questions, Wrong Ba Yung Nakablack Na Answer?????KC White Dela RosaNo ratings yet
- Corporate Governance Guidelines GMDocument15 pagesCorporate Governance Guidelines GMWaqas MahmoodNo ratings yet
- APPSC GR I Initial Key Paper IIDocument52 pagesAPPSC GR I Initial Key Paper IIdarimaduguNo ratings yet
- Topics For Oral PresentationDocument6 pagesTopics For Oral PresentationMohd HyqalNo ratings yet
- Gulf Case Study SolnDocument9 pagesGulf Case Study SolnHarsh SNo ratings yet
- Repro Indo China Conf PDFDocument16 pagesRepro Indo China Conf PDFPavit KaurNo ratings yet
- E34-1 Battery Charging and Dishcharging BoardDocument23 pagesE34-1 Battery Charging and Dishcharging BoardGanesa MurthyNo ratings yet
- Araldite 2020 + XW 396 - XW 397Document6 pagesAraldite 2020 + XW 396 - XW 397Paul Dan OctavianNo ratings yet
- Grammar and Oral Language Development (GOLD) : Reported By: Melyn A. Bacolcol Kate Batac Julie Ann OcampoDocument17 pagesGrammar and Oral Language Development (GOLD) : Reported By: Melyn A. Bacolcol Kate Batac Julie Ann Ocampoclara dupitasNo ratings yet
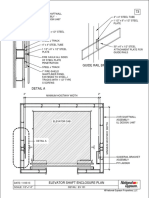
- Guide Rail Bracket AssemblyDocument1 pageGuide Rail Bracket AssemblyPrasanth VarrierNo ratings yet
- Shsa1105 - Unit-III Course MaterialsDocument58 pagesShsa1105 - Unit-III Course Materialssivanikesh bonagiriNo ratings yet
- Steinecker Boreas: Wort Stripping of The New GenerationDocument16 pagesSteinecker Boreas: Wort Stripping of The New GenerationAlejandro Javier Delgado AraujoNo ratings yet
- Jose André Morales, PH.D.: Ingeniería SocialDocument56 pagesJose André Morales, PH.D.: Ingeniería SocialJYMYNo ratings yet
- Topics For AssignmentDocument2 pagesTopics For AssignmentniharaNo ratings yet
- BMW Engine Wiring 2 OxDocument4 pagesBMW Engine Wiring 2 OxTomáš TdcNo ratings yet
- CP AssignmentDocument5 pagesCP AssignmentMSSM EngineeringNo ratings yet
- CHECK - Chapter 11 TCD AnswersDocument6 pagesCHECK - Chapter 11 TCD AnswersbonolomphaNo ratings yet
- Comparativa Microplex F40 Printronix P8220 enDocument1 pageComparativa Microplex F40 Printronix P8220 enangel ricaNo ratings yet
- English Literature Coursework Aqa GcseDocument6 pagesEnglish Literature Coursework Aqa Gcsef5d17e05100% (2)
- Case Study StarbucksDocument2 pagesCase Study StarbucksSonal Agarwal100% (2)
- What Is The Effective Method For Dissolution of HDPE and LDPE - PDFDocument12 pagesWhat Is The Effective Method For Dissolution of HDPE and LDPE - PDFAliNo ratings yet
- FoodhallDocument3 pagesFoodhallswopnilrohatgiNo ratings yet
- Acm Queue PDFDocument12 pagesAcm Queue PDFShubham Anil ShahareNo ratings yet
- InfltiDocument13 pagesInfltiLEKH021No ratings yet
- Material List Summary-WaptechDocument5 pagesMaterial List Summary-WaptechMarko AnticNo ratings yet
- TMIS07 - Kalam Internship - S7 Tesla MindsDocument3 pagesTMIS07 - Kalam Internship - S7 Tesla MindsDMJ JonesNo ratings yet