You might also like
- Greifeneder, Bless, Pham - 2011 - When Do People Rely On Affective and Cognitive Feelings in Judgment A ReviewDocument35 pagesGreifeneder, Bless, Pham - 2011 - When Do People Rely On Affective and Cognitive Feelings in Judgment A ReviewDave SainsburyNo ratings yet
- Berridge 2003 Parsing Reward, Paper Muy Bueno DopaminaDocument7 pagesBerridge 2003 Parsing Reward, Paper Muy Bueno DopaminaVissente TapiaNo ratings yet
- Scherer Moors 2018 Ann RevDocument58 pagesScherer Moors 2018 Ann RevPatricia ReyesNo ratings yet
- A Mixed-Method Approach On The Emotional Intelligence of Both Teachers and Students of Malvar School of Arts and Trade. Basis For An Enhanced Psychological and Counseling Program (EPCP)Document12 pagesA Mixed-Method Approach On The Emotional Intelligence of Both Teachers and Students of Malvar School of Arts and Trade. Basis For An Enhanced Psychological and Counseling Program (EPCP)Psychology and Education: A Multidisciplinary JournalNo ratings yet
- Causes and consequences of emotions on consumer behaviour: A review and integrative cognitive appraisal theoryDocument25 pagesCauses and consequences of emotions on consumer behaviour: A review and integrative cognitive appraisal theoryfarumasNo ratings yet
- Comparison of The Determinants For Positive and Negative Affect Proposed by Appraisal Theories, Goal-Directed Theories, and Predictive Processing TheoriesDocument6 pagesComparison of The Determinants For Positive and Negative Affect Proposed by Appraisal Theories, Goal-Directed Theories, and Predictive Processing TheoriesErin NasharuddinNo ratings yet
- JEduHealthPromot91157-4175279 113552Document8 pagesJEduHealthPromot91157-4175279 113552MCarmen MartinezNo ratings yet
- Two-Factor Theory of Emotion: Heart and Mind in HarmonyFrom EverandTwo-Factor Theory of Emotion: Heart and Mind in HarmonyNo ratings yet
- 1 s2.0 S0191886911003291 MainDocument6 pages1 s2.0 S0191886911003291 MainJesúsNo ratings yet
- Emotion and Motivation I: Defensive and Appetitive Reactions in Picture ProcessingDocument25 pagesEmotion and Motivation I: Defensive and Appetitive Reactions in Picture ProcessingfranciscoNo ratings yet
- Semantic Space TheoryDocument13 pagesSemantic Space TheoryMatteo LionelloNo ratings yet
- Greenberg, Emotion and CognitionDocument11 pagesGreenberg, Emotion and CognitionRamona FodorNo ratings yet
- Bagozzi, Gopinath, Nyer - 1999 - The Role of Emotions in MarketingDocument23 pagesBagozzi, Gopinath, Nyer - 1999 - The Role of Emotions in Marketingwext88No ratings yet
- The Role of Emotion in MarketingDocument23 pagesThe Role of Emotion in MarketingSandy StancuNo ratings yet
- The Role of Emotions in Marketing: Richard P. BagozziDocument23 pagesThe Role of Emotions in Marketing: Richard P. BagozziEngin OzsahinNo ratings yet
- Research Paper On EmotionsDocument8 pagesResearch Paper On Emotionskgtsnhrhf100% (1)
- Does Emotion Influence Visual Perception Depends On How You Look at ItDocument9 pagesDoes Emotion Influence Visual Perception Depends On How You Look at ItKatarina JovanovicNo ratings yet
- 2022 RyanDeci SDT EncyclopediaDocument7 pages2022 RyanDeci SDT EncyclopediauricomasNo ratings yet
- The EMO-Model: An Agent-Based Model of Primate Social Behavior Regulated by Two Emotional Dimensions, Anxiety-FEAR and Satisfaction-LIKEDocument25 pagesThe EMO-Model: An Agent-Based Model of Primate Social Behavior Regulated by Two Emotional Dimensions, Anxiety-FEAR and Satisfaction-LIKEJuan TobarNo ratings yet
- Metacognition, Emotional Intelligence, Aggression and Passionate Love Among WomenDocument129 pagesMetacognition, Emotional Intelligence, Aggression and Passionate Love Among WomenjoseNo ratings yet
- Motivation Paper 2Document15 pagesMotivation Paper 2mabney75No ratings yet
- The Control-Value Theory of Achievement Emotions: Assumptions, Corollaries, and Implications For Educational Research and PracticeDocument27 pagesThe Control-Value Theory of Achievement Emotions: Assumptions, Corollaries, and Implications For Educational Research and PracticeBayarmaa BuyantogtokhNo ratings yet
- NIH Public Access: Author ManuscriptDocument43 pagesNIH Public Access: Author ManuscriptAntónio MartinsNo ratings yet
- Emotional Intelligence Effects in the WorkplaceDocument24 pagesEmotional Intelligence Effects in the WorkplaceAnonymous 6rn7803No ratings yet
- Positive Emotions in EducationDocument15 pagesPositive Emotions in Educationmanuel alvarezNo ratings yet
- Human BehaviorDocument6 pagesHuman BehaviorsurjitxinghaNo ratings yet
- AffectDocument6 pagesAffectVeer SinghNo ratings yet
- I.E. They Are Affected By, But Do Not Affect, The Decision ProcessDocument4 pagesI.E. They Are Affected By, But Do Not Affect, The Decision ProcessCarl HartfordNo ratings yet
- Emotions Research PaperDocument8 pagesEmotions Research Paperafedsdlmz100% (1)
- Happiness, Personality Traits Predict College Teacher OptimismDocument18 pagesHappiness, Personality Traits Predict College Teacher OptimismSamia GhafoorNo ratings yet
- PRE 705 Week 7 DiscussionDocument3 pagesPRE 705 Week 7 DiscussionsarahthamNo ratings yet
- Incentive Theory and Workplace MotivationDocument7 pagesIncentive Theory and Workplace MotivationNIMRAH RAISNo ratings yet
- The Psychological Flexibility Questionnaire (PFQ) : Development, Reliability and ValidityDocument10 pagesThe Psychological Flexibility Questionnaire (PFQ) : Development, Reliability and ValidityMónika MikeNo ratings yet
- The Role of Emotions in MarketingDocument23 pagesThe Role of Emotions in Marketingluizamj100% (5)
- The Broaden-And-build Theory of Positive Emotions - BL Frederickson - Royal SocietyDocument11 pagesThe Broaden-And-build Theory of Positive Emotions - BL Frederickson - Royal Societyswetava_ganguliNo ratings yet
- 3 Lazarus 1988Document12 pages3 Lazarus 1988claudia osorioNo ratings yet
- Behavioral Sciences: The Basic Psychological Needs in The Classroom Scale (BPN-CS)Document13 pagesBehavioral Sciences: The Basic Psychological Needs in The Classroom Scale (BPN-CS)Srishti GaurNo ratings yet
- Acceptance Alone Is A Better Predictor of Psychopathology and Well-Being Than Emotional Competence, Emotion Regulation and MindfulnessDocument4 pagesAcceptance Alone Is A Better Predictor of Psychopathology and Well-Being Than Emotional Competence, Emotion Regulation and Mindfulnesstimsmith1081574No ratings yet
- SE Informed Group Psychotherapy For Intl Journal of Group PsychotherapyDocument13 pagesSE Informed Group Psychotherapy For Intl Journal of Group PsychotherapyLella Misikir100% (1)
- A Study of The Relation Between Moral Judgment and Emotional IntelligenceDocument8 pagesA Study of The Relation Between Moral Judgment and Emotional Intelligenceella003No ratings yet
- Emotion Regulation in ACT - Steven C. HayesDocument13 pagesEmotion Regulation in ACT - Steven C. HayesVartax100% (2)
- Social Emotions Emotions in Social InterDocument7 pagesSocial Emotions Emotions in Social InterDunja Chrysina ChrysargyreaNo ratings yet
- The Neurodevelopment of Empathy in HumanDocument11 pagesThe Neurodevelopment of Empathy in HumanMadina KhasanboevaNo ratings yet
- Personality and Individual Di Fferences: David Preece, Rodrigo Becerra, Ken Robinson, Justine Dandy, Alfred AllanDocument13 pagesPersonality and Individual Di Fferences: David Preece, Rodrigo Becerra, Ken Robinson, Justine Dandy, Alfred Allanmartina.giuffrida847No ratings yet
- 2022 - Silva Et AlDocument20 pages2022 - Silva Et AlMarta MatosNo ratings yet
- 2012_A framework for emergent emotions, based on motivation and cognitive modulatorsDocument24 pages2012_A framework for emergent emotions, based on motivation and cognitive modulatorsvincent.wienerroither1287No ratings yet
- Appraisal Theory of Emotion IntroductionDocument10 pagesAppraisal Theory of Emotion IntroductionAndra ToldeanNo ratings yet
- NC_ED_in_2007_DamasioDocument8 pagesNC_ED_in_2007_DamasioSANDRA ESTEFANÍA RIVERANo ratings yet
- Toward An Integrative Psychometric Model of EmotionsDocument25 pagesToward An Integrative Psychometric Model of EmotionsSandraNo ratings yet
- Research Paper EmotionsDocument4 pagesResearch Paper EmotionsafeawckewNo ratings yet
- Bonsa PsychologyDocument13 pagesBonsa PsychologybeekiijNo ratings yet
- PSY4111Document3 pagesPSY4111vylouisevanNo ratings yet
- 4 Heyes 2018 EmpathyDocument9 pages4 Heyes 2018 EmpathyRenata AndradeNo ratings yet
- Gale Researcher Guide for: Value- and Incentive-Theories of MotivationFrom EverandGale Researcher Guide for: Value- and Incentive-Theories of MotivationNo ratings yet
- Wright 2009Document13 pagesWright 20092332801101No ratings yet
- Positive PsychologyDocument3 pagesPositive PsychologySAKSHI GANDHINo ratings yet
- mainDocument10 pagesmainbonnie.a.omalleyNo ratings yet
- Carroll e Izard The Structure and Person PDFDocument37 pagesCarroll e Izard The Structure and Person PDFErick SuasteNo ratings yet
- The Appreciative Heart: The Psychophysiology of Positive Emotions and Optimal FunctioningDocument23 pagesThe Appreciative Heart: The Psychophysiology of Positive Emotions and Optimal FunctioningProfessor João GabrielNo ratings yet
- Theory of FunctionalismDocument2 pagesTheory of FunctionalismPoovinaa SeluanaikamNo ratings yet
- Eleanor Richardson, Jeremy S Lewis, Jo Gibson, Chris Morgan, Mark Halaki, Karen Ginn, Gillian YeowellDocument12 pagesEleanor Richardson, Jeremy S Lewis, Jo Gibson, Chris Morgan, Mark Halaki, Karen Ginn, Gillian YeowellVizaNo ratings yet
- Appendix: A Best Practices Resource Guide For Physical TherapistsDocument28 pagesAppendix: A Best Practices Resource Guide For Physical TherapistsVizaNo ratings yet
- SarabonEtAl AssessmentofIsometricTrunkStrength 2014Document7 pagesSarabonEtAl AssessmentofIsometricTrunkStrength 2014VizaNo ratings yet
- Fitness Testing Record 1204Document1 pageFitness Testing Record 1204VizaNo ratings yet
- Case Report Example and AbstractDocument8 pagesCase Report Example and AbstractMyra Lyn OloresNo ratings yet
- Sensorimotor Training A Global Approach For BalancDocument9 pagesSensorimotor Training A Global Approach For BalancVizaNo ratings yet
- A Pragmatic Regional Interdependence Approach To Primary Frozen Shoulder A Retrospective Case SeriesDocument11 pagesA Pragmatic Regional Interdependence Approach To Primary Frozen Shoulder A Retrospective Case SeriesVizaNo ratings yet
- HiDocument38 pagesHiNajeebuddin AhmedNo ratings yet
- Lumbar Extension ExercisesDocument3 pagesLumbar Extension ExercisessathishrajsNo ratings yet
- Endurance of Low Back Musculature: Normative Data For AdultsDocument10 pagesEndurance of Low Back Musculature: Normative Data For AdultsVizaNo ratings yet
- Diagnosis, Classification Management of Chronic Low Back PainDocument31 pagesDiagnosis, Classification Management of Chronic Low Back PainVizaNo ratings yet
- Measuring Clinical Reasoning and Interprofessional AttitudesDocument6 pagesMeasuring Clinical Reasoning and Interprofessional AttitudesVizaNo ratings yet
- Development of A Clinical Examination in Non-Specific Low Back Pain: A Delphi TechniqueDocument5 pagesDevelopment of A Clinical Examination in Non-Specific Low Back Pain: A Delphi TechniqueVizaNo ratings yet
- Pain Chronification What Should A Non Pain Medicine Specialist KnowDocument11 pagesPain Chronification What Should A Non Pain Medicine Specialist KnowVizaNo ratings yet
- Schoenfeld Volumen ReviewDocument6 pagesSchoenfeld Volumen ReviewVizaNo ratings yet
- BMC Musculoskeletal Disorders: Chronic Non-Specific Low Back Pain - Sub-Groups or A Single Mechanism?Document15 pagesBMC Musculoskeletal Disorders: Chronic Non-Specific Low Back Pain - Sub-Groups or A Single Mechanism?VizaNo ratings yet
- ChatDocument11 pagesChatVizaNo ratings yet
- Manual Physical Therapy For Chronic Pain The Complex Whole Is Greater Than The Sum of Its PartsDocument4 pagesManual Physical Therapy For Chronic Pain The Complex Whole Is Greater Than The Sum of Its PartsVizaNo ratings yet
- Patient Specific Functional ScaleDocument1 pagePatient Specific Functional Scalepfi_jenNo ratings yet
- Fatigue Severity ScaleDocument1 pageFatigue Severity ScaleAffuadiNo ratings yet
- Sullivan LaikysenaDocument9 pagesSullivan LaikysenaVizaNo ratings yet
- 00029Document11 pages00029rudhras22No ratings yet
- Exercise Induced Hypoalgesia After Acute And.11-2Document22 pagesExercise Induced Hypoalgesia After Acute And.11-2VizaNo ratings yet
- 10.1016@S0140 67361160610 7Document10 pages10.1016@S0140 67361160610 7Agus PrimaNo ratings yet
- Sacroiliac Joint Motion in Patients With Degenerative Lumbar Spine DisordersDocument8 pagesSacroiliac Joint Motion in Patients With Degenerative Lumbar Spine DisordersVizaNo ratings yet
- Effect of Graded Plank Protocol On Core Stability in Sedentary DentistsDocument5 pagesEffect of Graded Plank Protocol On Core Stability in Sedentary DentistsVizaNo ratings yet
- Jospt 2019 8248Document13 pagesJospt 2019 8248VizaNo ratings yet
- Effect of Graded Plank Protocol On Core Stability in Sedentary DentistsDocument5 pagesEffect of Graded Plank Protocol On Core Stability in Sedentary DentistsVizaNo ratings yet
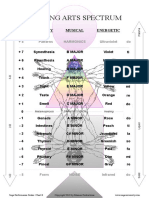
- Healing Arts Spectrum: Sensory Musical EnergeticDocument1 pageHealing Arts Spectrum: Sensory Musical EnergeticVizaNo ratings yet
- 7.17 Degenerative Disease of Hip Joint. Radiological DiagnosisDocument76 pages7.17 Degenerative Disease of Hip Joint. Radiological DiagnosisVizaNo ratings yet
- Gamification and Game Based LearningDocument6 pagesGamification and Game Based LearningSheryl Tubino EstacioNo ratings yet
- Creativity & Entrepreneurship GuideDocument6 pagesCreativity & Entrepreneurship Guideanika bhuianNo ratings yet
- Entropy, Creativity & Learning de LangeDocument282 pagesEntropy, Creativity & Learning de LangeGavin RitzNo ratings yet
- Hard Soft Emotional SkillsDocument42 pagesHard Soft Emotional SkillsImad AznagNo ratings yet
- Learning Guide Unit 1Document8 pagesLearning Guide Unit 1new workNo ratings yet
- Explaining How Leaders' Personal Traits Impact Foreign PolicyDocument41 pagesExplaining How Leaders' Personal Traits Impact Foreign PolicyEdson Neves Jr.100% (1)
- Unit-V Consumer Behaviour (CB) : Meaning and SignificanceDocument19 pagesUnit-V Consumer Behaviour (CB) : Meaning and Significancemahatheer mohamedNo ratings yet
- 3228 ShaharyrDocument40 pages3228 ShaharyrAwais AkramNo ratings yet
- Crosshead Section PresentationDocument42 pagesCrosshead Section PresentationGio LagadiaNo ratings yet
- The Impact of Motivation On Reading ComprehensionDocument7 pagesThe Impact of Motivation On Reading ComprehensionDao NguyenNo ratings yet
- OraDocument31 pagesOraAndrew OrañoNo ratings yet
- The Effects of Watching TV Cartoon Program On The Behavior of Grade One Pupils in Brion-Silva Elementary SchoolDocument34 pagesThe Effects of Watching TV Cartoon Program On The Behavior of Grade One Pupils in Brion-Silva Elementary SchoolMelannie D Arcenas100% (2)
- NVFC Arson ReportDocument36 pagesNVFC Arson ReportepraetorianNo ratings yet
- Empathize Through Observation and InterviewsDocument10 pagesEmpathize Through Observation and InterviewsPhilosofee HijabNo ratings yet
- AGNEW 2016 CriminologyDocument31 pagesAGNEW 2016 CriminologyRafael CarvalhoNo ratings yet
- HCI Persuasive Technology PresentationDocument27 pagesHCI Persuasive Technology PresentationCahyo Al- HazimNo ratings yet
- Leadership Styles & TacticsDocument29 pagesLeadership Styles & TacticsNISHA100% (40)
- Appraisal ScriptDocument14 pagesAppraisal ScriptSai KrishnaNo ratings yet
- Toaz - Info Personal Development DLL PRDocument45 pagesToaz - Info Personal Development DLL PRArcee DascoNo ratings yet
- Nemo Dat Quod Non Habet The Lived Experi 635de2fbDocument11 pagesNemo Dat Quod Non Habet The Lived Experi 635de2fbTabuk City 098No ratings yet
- Entrep - Unit - I CorrectedDocument2,677 pagesEntrep - Unit - I CorrectedJayati BohreNo ratings yet
- AFFECTIVE TARGETS: AttitudeDocument19 pagesAFFECTIVE TARGETS: AttitudeJaika SedoroNo ratings yet
- Peer Tutoring For Math PDFDocument8 pagesPeer Tutoring For Math PDFJean Maquilan BatoNo ratings yet
- Performance Management Process Torrington and Hall Model PDFDocument16 pagesPerformance Management Process Torrington and Hall Model PDFKiran Prasannan Sheela67% (3)
- The Study of University Students' Motivation: Jana M. Safranková, Martin SikyrDocument12 pagesThe Study of University Students' Motivation: Jana M. Safranková, Martin SikyrCharito BedaniaNo ratings yet
- The Relationship of Principal Leadership Behaviors With School Climate, Teacher Job Satisfaction, and Student AchievementDocument133 pagesThe Relationship of Principal Leadership Behaviors With School Climate, Teacher Job Satisfaction, and Student AchievementNuraisyah NordinNo ratings yet
- Humanistic ApproachDocument20 pagesHumanistic ApproachNur FarahinNo ratings yet
- Anthony Robbins Action BookDocument27 pagesAnthony Robbins Action Booknahsang100% (19)
- Chapter 4Document40 pagesChapter 4Roopa YadavNo ratings yet
- PSY 100-Human Behavior-Uzma Mazhar-Yasser Hashmi-Zahbia Sarfraz PDFDocument3 pagesPSY 100-Human Behavior-Uzma Mazhar-Yasser Hashmi-Zahbia Sarfraz PDFMinhal MalikNo ratings yet