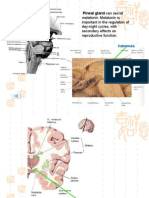
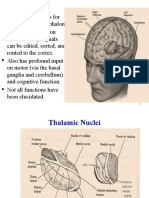
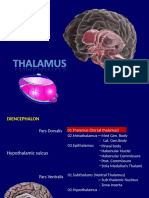
You might also like
- El Tálamo: Aspectos Neurofuncionales: Related PapersDocument8 pagesEl Tálamo: Aspectos Neurofuncionales: Related PapersAlvear KatitaNo ratings yet
- Tálamo y FuncionesDocument13 pagesTálamo y FuncionesMaría Fernanda OréNo ratings yet
- TalamoDocument7 pagesTalamoDanya RoblesNo ratings yet
- The Thalamus: A Review of Its Functional Anatomy: AuthorDocument9 pagesThe Thalamus: A Review of Its Functional Anatomy: Authorranu anggaraNo ratings yet
- Course 3.1 Diencephalon 3 L 6 THALAMUSDocument74 pagesCourse 3.1 Diencephalon 3 L 6 THALAMUSSuman MahmoodNo ratings yet
- Thalamus: What Is The Thalamus and What Are Its Main Functions?Document8 pagesThalamus: What Is The Thalamus and What Are Its Main Functions?sandeep1699No ratings yet
- An Introduction To Neural SystemsDocument41 pagesAn Introduction To Neural SystemsFebyNo ratings yet
- Thalamus Structure Relays Sensory InfoDocument7 pagesThalamus Structure Relays Sensory InfoYubi KimNo ratings yet
- Basal GangliaDocument4 pagesBasal GangliaOvolime UbodiomNo ratings yet
- L20-Thalamus & Limbic SystemDocument33 pagesL20-Thalamus & Limbic SystemSk JsNo ratings yet
- Fundamentals of The Nervous SystemDocument34 pagesFundamentals of The Nervous SystemEthan Dreiz BaltazarNo ratings yet
- Catatan Dokter Muda Anatomy OtakDocument23 pagesCatatan Dokter Muda Anatomy OtakFerdi StefiyanNo ratings yet
- Diencephalon Structure and FunctionsDocument29 pagesDiencephalon Structure and FunctionsmarciaNo ratings yet
- Topic 2: The Extrapyramidal System. The CerebellumDocument42 pagesTopic 2: The Extrapyramidal System. The Cerebellumprashant singhNo ratings yet
- Thalamus 130624023852 Phpapp02Document37 pagesThalamus 130624023852 Phpapp02Dr Abdulkadir Aminu DaudaNo ratings yet
- Thalamus: - Dr. Sachin A. AdukiaDocument56 pagesThalamus: - Dr. Sachin A. AdukiaNairda TorresNo ratings yet
- Cerebellum & DiencephalonDocument21 pagesCerebellum & Diencephalonprasun_vNo ratings yet
- Anatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessFrom EverandAnatomy of Neuropsychiatry: The New Anatomy of the Basal Forebrain and Its Implications for Neuropsychiatric IllnessNo ratings yet
- ThalamusDocument21 pagesThalamusTem GemechuNo ratings yet
- 5.basal Ganglia-2016 - 082539Document28 pages5.basal Ganglia-2016 - 082539sirima makuraNo ratings yet
- Thalamic Neuromodulation and Its Implications For Executive NetworksDocument22 pagesThalamic Neuromodulation and Its Implications For Executive NetworkstratrNo ratings yet
- Dorsal Part of The DiencephalonDocument7 pagesDorsal Part of The DiencephalonThe Panda EntertainerNo ratings yet
- 6 Diencephalon 2022Document51 pages6 Diencephalon 2022Gregory WigginsNo ratings yet
- Thalamus NotesDocument11 pagesThalamus NotesTanvir AhmedNo ratings yet
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesFrom EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNo ratings yet
- The MidbrainDocument3 pagesThe MidbrainAshly Kate AbarientosNo ratings yet
- Snell Reticular Formation and Limbic SystemDocument9 pagesSnell Reticular Formation and Limbic SystemFrnz RiveraNo ratings yet
- UntitledDocument5 pagesUntitledRafu DestajoNo ratings yet
- Basic Basal Ganglia: The Basal Ganglia Consist of Several Interconnected NucleiDocument5 pagesBasic Basal Ganglia: The Basal Ganglia Consist of Several Interconnected NucleiFakhriNo ratings yet
- Histology of The CNSDocument7 pagesHistology of The CNStiiandiNo ratings yet
- ThalamusDocument30 pagesThalamusRho Vince Caño Malagueño100% (1)
- ThalamusDocument17 pagesThalamusPrafulla KasarNo ratings yet
- 18 - Thalamus and Limbic System (Edited)Document22 pages18 - Thalamus and Limbic System (Edited)Fotocopias LulisNo ratings yet
- The Brain: Functional Divisions: Primary and Secondary Vesicles of The Developing CNSDocument6 pagesThe Brain: Functional Divisions: Primary and Secondary Vesicles of The Developing CNSKelly FariaNo ratings yet
- Adrenal GlandDocument7 pagesAdrenal GlandAbdullah Yeamin PeyalNo ratings yet
- Mind at Rest: How Neuron Structure Evolves in the Sleep Cycle.From EverandMind at Rest: How Neuron Structure Evolves in the Sleep Cycle.No ratings yet
- Nervous System Functions and Neuron Anatomy ExplainedDocument184 pagesNervous System Functions and Neuron Anatomy ExplainedAbdurahman TainyNo ratings yet
- Case 6 Revision - Cognitive Impairment: Central SulcusDocument7 pagesCase 6 Revision - Cognitive Impairment: Central SulcusCharlie WalkerNo ratings yet
- 116 - Concept of Perception and CoordinationDocument3 pages116 - Concept of Perception and Coordinationalaisahmae02No ratings yet
- Human BrainDocument44 pagesHuman BrainSri vaishnava100% (1)
- Fisiologia Del Sistema Nervioso AutonomoDocument11 pagesFisiologia Del Sistema Nervioso Autonomodavid bellidoNo ratings yet
- Circuitos Limbico-TalamicoDocument50 pagesCircuitos Limbico-TalamicomarielaNo ratings yet
- Anatomy and Physiology of the ThalamusDocument11 pagesAnatomy and Physiology of the ThalamusMayur ShindeNo ratings yet
- Arterial Supply of The Thalamus - A Comprehensive ReviewDocument9 pagesArterial Supply of The Thalamus - A Comprehensive Reviewمحمد رؤوف خير الدينNo ratings yet
- In NeuroanatomyDocument42 pagesIn NeuroanatomycristieristiieNo ratings yet
- Basal GangliaDocument9 pagesBasal GangliaKatherine NunnNo ratings yet
- NByNF 2Document29 pagesNByNF 2Oriana CofréNo ratings yet
- Frontocoritical CircuitsDocument29 pagesFrontocoritical CircuitsEmmanuel Domínguez RosalesNo ratings yet
- Thalamus Abd MajidDocument25 pagesThalamus Abd MajidKAKANYERO SUNDAY ROBERTNo ratings yet
- What Are The Basal Ganglia?Document53 pagesWhat Are The Basal Ganglia?Gursangeetkaur2010No ratings yet
- Basal GangliaDocument11 pagesBasal GangliaSTEVEN OKURUTNo ratings yet
- Week 1 Nervous TissueDocument29 pagesWeek 1 Nervous TissueArunesh 456No ratings yet
- Nervous System Anatomy and FunctionsDocument31 pagesNervous System Anatomy and FunctionsSalem AlshahraniNo ratings yet
- The Thalamus Is A Scanner: All Information Comes Into The Thalamus Before Being Distributed To Other Parts of The BrainDocument54 pagesThe Thalamus Is A Scanner: All Information Comes Into The Thalamus Before Being Distributed To Other Parts of The BrainBaguma MichaelNo ratings yet
- Brain NeurotransmittersDocument29 pagesBrain Neurotransmittersmaha abdallahNo ratings yet
- Homeostasis and Higher Brain FunctionsDocument84 pagesHomeostasis and Higher Brain Functionsbodeadumitru9261No ratings yet
- Cerebellum Anatomy and ConnectionsDocument36 pagesCerebellum Anatomy and ConnectionssaskiabpNo ratings yet
- Fisiologia Del Sistema Nervioso AutonomoDocument11 pagesFisiologia Del Sistema Nervioso AutonomoMerci BaezNo ratings yet
- BioPsychology Chapter 34Document10 pagesBioPsychology Chapter 34Johnreih BanggaNo ratings yet
- p4. ThalamusDocument26 pagesp4. ThalamusGedefaw TegegneNo ratings yet
- Are Subjective Cognitive Complaints A Risk Factor For DementiaDocument10 pagesAre Subjective Cognitive Complaints A Risk Factor For DementiaFelipeWebster CorderoNo ratings yet
- Attention and Executive Control Predict AlzDocument8 pagesAttention and Executive Control Predict AlzFelipeWebster CorderoNo ratings yet
- Cognitive and Adaptive Outcome Un Low-Grade Pediatric Cerebelar AstrocytomasDocument7 pagesCognitive and Adaptive Outcome Un Low-Grade Pediatric Cerebelar AstrocytomasFelipeWebster CorderoNo ratings yet
- Estructura y Funcion Del CerebeloDocument8 pagesEstructura y Funcion Del Cerebeloneuropat100% (1)
- Cerebelo y CognicionDocument11 pagesCerebelo y CognicionFelipeWebster CorderoNo ratings yet
- ACE Add en Brooke AltaeducacionDocument5 pagesACE Add en Brooke AltaeducacionClaudio Fernando Medina HernandezNo ratings yet
- Cerebellum Ang CognitionDocument14 pagesCerebellum Ang CognitionRenata ZucarinoNo ratings yet
- Incidencia Clinica Demencia Frontal LeidoDocument7 pagesIncidencia Clinica Demencia Frontal LeidoFelipeWebster CorderoNo ratings yet
- FAB Spanish Application of Frontal Assessment BatteryDocument5 pagesFAB Spanish Application of Frontal Assessment BatteryFelipeWebster CorderoNo ratings yet
- Marketing Management - Module 7Document18 pagesMarketing Management - Module 7SnehaNo ratings yet
- DashaDocument2 pagesDashaSrini VasanNo ratings yet
- Graduate School: Mariano Marcos State UniversityDocument2 pagesGraduate School: Mariano Marcos State UniversityLeslie Anne BiteNo ratings yet
- Brian Leggett (Ed) 2006 Rhetorical-Approach-To-PersuasionDocument9 pagesBrian Leggett (Ed) 2006 Rhetorical-Approach-To-PersuasionM MoralesNo ratings yet
- Dosi ResDocument2 pagesDosi Resapi-270166579No ratings yet
- Test Specification: iUBT435 - Anatomy and PhysiologyDocument3 pagesTest Specification: iUBT435 - Anatomy and PhysiologyVicki TseNo ratings yet
- Demystifying Interventional Radiology A Guide For Medical StudentsDocument195 pagesDemystifying Interventional Radiology A Guide For Medical StudentsMo Haroon100% (1)
- Test 2: Section 1Document5 pagesTest 2: Section 1Wall HarryNo ratings yet
- CorpusRembrandt 3Document819 pagesCorpusRembrandt 3Yuliya UdotNo ratings yet
- Early Childhood Physical DevelopmentDocument34 pagesEarly Childhood Physical DevelopmentEditha100% (1)
- Visual Division-1Document3 pagesVisual Division-1Jumran BaharNo ratings yet
- Manual Book Verticality by TBiGMobileDocument27 pagesManual Book Verticality by TBiGMobileMuhammad Maulana SetiawanNo ratings yet
- Dual Nature of Radiation and Matter PDFDocument18 pagesDual Nature of Radiation and Matter PDFLakshit SharmaNo ratings yet
- Ded Moroz Minitheme by SlidesgoDocument41 pagesDed Moroz Minitheme by SlidesgoTazkia AuditaNo ratings yet
- Watching A Typhoon Using Sentinel-1Document39 pagesWatching A Typhoon Using Sentinel-1James GulreyNo ratings yet
- CHEM 1211K Lab Fall 2020: Submission GuideDocument3 pagesCHEM 1211K Lab Fall 2020: Submission GuideParker Santo DomingoNo ratings yet
- DAY 1 Passage 1Document2 pagesDAY 1 Passage 1wick90715No ratings yet
- Effect of Summer Green Gram (Vigna Radiata L.) Varieties, Sulphur Levels and Fertilizer Levels On Quality, Nutrient Content and Uptake Under South Gujarat ConditionDocument3 pagesEffect of Summer Green Gram (Vigna Radiata L.) Varieties, Sulphur Levels and Fertilizer Levels On Quality, Nutrient Content and Uptake Under South Gujarat ConditionThakor BharviNo ratings yet
- Numerology Decoder PDFDocument3 pagesNumerology Decoder PDFChris Walker100% (3)
- IB PHYSICS 2.4 Presentation All UnitDocument34 pagesIB PHYSICS 2.4 Presentation All UnitJustCallMeLarryNo ratings yet
- Stress Distribution in The Temporomandibular Joint After Mandibular Protraction A Three-Dimensional Finite Element StudyDocument10 pagesStress Distribution in The Temporomandibular Joint After Mandibular Protraction A Three-Dimensional Finite Element StudySelvaNo ratings yet
- CMC 63rd Batch Class Routine for Thorax CardDocument5 pagesCMC 63rd Batch Class Routine for Thorax CardMath NewNo ratings yet
- Greater Noida HousingDocument42 pagesGreater Noida HousingYathaarth RastogiNo ratings yet
- Chapter 15-Pipe Drains PDFDocument33 pagesChapter 15-Pipe Drains PDFjpmega004524No ratings yet
- Attractive Celebrity and Peer Images On Instagram. Effect On Women's Mood and Body ImageDocument7 pagesAttractive Celebrity and Peer Images On Instagram. Effect On Women's Mood and Body ImageDesiré Abrante RodríguezNo ratings yet
- Auveco HNI 490Document1 pageAuveco HNI 490BillNo ratings yet
- Grade 4 DLL Quarter 2 Week 9 (Sir Bien Cruz)Document38 pagesGrade 4 DLL Quarter 2 Week 9 (Sir Bien Cruz)FUMIKO SOPHIANo ratings yet
- MTech Seminar II and Project SyllabusDocument3 pagesMTech Seminar II and Project SyllabusrcherrNo ratings yet
- DRRMQ 3 W1Document44 pagesDRRMQ 3 W1Edelmar BenosaNo ratings yet
- CASE-STUDY of NFPTs in Butuan CityDocument35 pagesCASE-STUDY of NFPTs in Butuan CityKizza Eyn MinguitaNo ratings yet