You might also like
- Trefoil Factor 2 Promotes Cell Proliferation in Pancreatic - Cells Through CXCR-4-Mediated ERK1/2 PhosphorylationDocument11 pagesTrefoil Factor 2 Promotes Cell Proliferation in Pancreatic - Cells Through CXCR-4-Mediated ERK1/2 PhosphorylationArshad AliNo ratings yet
- ZBC 23307Document15 pagesZBC 23307Michael UNo ratings yet
- Prostaglandin Receptor SignallingDocument7 pagesProstaglandin Receptor SignallingAdair BlasNo ratings yet
- 03JBC-Bae-amniotic FluidDocument6 pages03JBC-Bae-amniotic Fluid이용문No ratings yet
- Biolreprod 0696Document11 pagesBiolreprod 0696Ahmad SolihinNo ratings yet
- A Postgenomic Integratred View of Prostaglandins in Reproduction: Implications For Other Body SystemsDocument25 pagesA Postgenomic Integratred View of Prostaglandins in Reproduction: Implications For Other Body SystemsNabila FNo ratings yet
- Ikemoto 2002Document13 pagesIkemoto 2002cataawwwNo ratings yet
- REGgamma and Chromasomal StabiligyDocument37 pagesREGgamma and Chromasomal Stabiligym9bpb6frpxNo ratings yet
- Park 2003Document8 pagesPark 2003lisnallNo ratings yet
- The Emerging Physiological Roles of The Glycerophosphodiesterase FamilyDocument19 pagesThe Emerging Physiological Roles of The Glycerophosphodiesterase FamilyArshia NazirNo ratings yet
- Cyclooxygenase-2 Biology: Joan ClàriaDocument15 pagesCyclooxygenase-2 Biology: Joan ClàriajustrudinNo ratings yet
- Rho-Kinase Regulates Prostaglandin D - Stimulated Heat Shock Protein 27 Induction in OsteoblastsDocument5 pagesRho-Kinase Regulates Prostaglandin D - Stimulated Heat Shock Protein 27 Induction in OsteoblastsIsaac GoldNo ratings yet
- J. Biol. Chem.-2002-Ma-39343-9Document8 pagesJ. Biol. Chem.-2002-Ma-39343-9Melin YohanaNo ratings yet
- Glutamine's role in neutrophil superoxide production and NADPH oxidase component expressionDocument6 pagesGlutamine's role in neutrophil superoxide production and NADPH oxidase component expressionAstynnia GanNo ratings yet
- Signaling Via Macrophage G2A Enhances Efferocytosis of Dying Neutrophils by Augmentation of Rac ActivityDocument15 pagesSignaling Via Macrophage G2A Enhances Efferocytosis of Dying Neutrophils by Augmentation of Rac ActivityTheodora TeddyNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument6 pagesBiochemical and Biophysical Research CommunicationsSebastian LopezNo ratings yet
- J. Biol. Chem.-2000-Del Peso-27205-11Document7 pagesJ. Biol. Chem.-2000-Del Peso-27205-11Thomas IngallsNo ratings yet
- Intracellular Glycerol Influences Resistance To Freeze Stress in Saccharomyces Cerevisiae Analysis of A Quadruple Mutant in Glycerol Dehydrogenase Genes and Glycerol-Enriched Cells 2004Document7 pagesIntracellular Glycerol Influences Resistance To Freeze Stress in Saccharomyces Cerevisiae Analysis of A Quadruple Mutant in Glycerol Dehydrogenase Genes and Glycerol-Enriched Cells 2004Abo Ahmad AlsaidiNo ratings yet
- Prostaglandin E Synthesis and Secretion: The Role of PGE SynthasesDocument12 pagesProstaglandin E Synthesis and Secretion: The Role of PGE SynthasesKarlaNo ratings yet
- The Basis of Prostaglandin Synthesis in CoralDocument9 pagesThe Basis of Prostaglandin Synthesis in CoralaliNo ratings yet
- Prostaglandin: Continued DonationsDocument27 pagesProstaglandin: Continued DonationsrajeevmkNo ratings yet
- Requirement of Vesicle-Associated Membrane Protein 721 and 722 For Sustained Growth During Immune Responses in ArabidopsisDocument8 pagesRequirement of Vesicle-Associated Membrane Protein 721 and 722 For Sustained Growth During Immune Responses in ArabidopsisAlberto MachoNo ratings yet
- Caspase Inhibitor Diminishes Caffeic Acid-Induced Apoptosis in Osteosarcoma CellsDocument5 pagesCaspase Inhibitor Diminishes Caffeic Acid-Induced Apoptosis in Osteosarcoma CellsMalik AkbarNo ratings yet
- Fenech 2020Document7 pagesFenech 2020Melissa SouzaNo ratings yet
- 2-Hydroxyestradiol Attenuates Renal Disease in Chronic Puromycin Aminonucleoside NephropathyDocument11 pages2-Hydroxyestradiol Attenuates Renal Disease in Chronic Puromycin Aminonucleoside NephropathyFauziana NurhanisahNo ratings yet
- A Postgenomic Integratred View of Prostaglandins IDocument26 pagesA Postgenomic Integratred View of Prostaglandins InaufalNo ratings yet
- The Genetic Aspect and Morphological Appearance of AchondrogenesisDocument10 pagesThe Genetic Aspect and Morphological Appearance of AchondrogenesisRizka RamadaniNo ratings yet
- MainDocument8 pagesMainAry MadinaNo ratings yet
- 2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Document9 pages2014-Echeverria C-Endotoxin-induced endothelial fibrosis is dependent on the expression of TGF-β1 and TGF-β2Pablo Tapia BatallaNo ratings yet
- Atopic Dermatitis and Filaggrin: SciencedirectDocument8 pagesAtopic Dermatitis and Filaggrin: SciencedirectMisaeldpdNo ratings yet
- Atopic Dermatitis and Filaggrin: SciencedirectDocument8 pagesAtopic Dermatitis and Filaggrin: SciencedirectMisaeldpdNo ratings yet
- 1 s2.0 S0002944010648523 MainDocument8 pages1 s2.0 S0002944010648523 MaindpcamposhNo ratings yet
- Dapagliflozin Binds Specifically To Sodium-Glucose Cotransporter 2 in The Proximal Renal Tubule - PMCDocument19 pagesDapagliflozin Binds Specifically To Sodium-Glucose Cotransporter 2 in The Proximal Renal Tubule - PMCBunny SmithNo ratings yet
- GM3 GangliosideDocument6 pagesGM3 Gangliosideanon_818654990No ratings yet
- Fullerenes As An Effective Amyloid Fibrils Disaggregating Nanomaterialibqvb PDFDocument2 pagesFullerenes As An Effective Amyloid Fibrils Disaggregating Nanomaterialibqvb PDFjeepdime3No ratings yet
- 2019 Vaamonde-GarciaDocument13 pages2019 Vaamonde-GarciaHistologia elementarNo ratings yet
- Cyclic AMP and Glycogen Synthase Kinase 3 Form A RegulatoryDocument14 pagesCyclic AMP and Glycogen Synthase Kinase 3 Form A RegulatorySouvik DeyNo ratings yet
- Tugas Baca 3,4Document6 pagesTugas Baca 3,4Jina KimNo ratings yet
- Galactosidase Over-Production by A Mig1 Mutant of Kluyveromyces Marxianus KM For Efficient Hydrolysis of LactoseDocument8 pagesGalactosidase Over-Production by A Mig1 Mutant of Kluyveromyces Marxianus KM For Efficient Hydrolysis of Lactose1039danielNo ratings yet
- (14796813 - Journal of Molecular Endocrinology) Larval Zebrafish As A Model For Glucose Metabolism - Expression of Phosphoenolpyruvate Carboxykinase As A Marker For Exposure To Anti-Diabetic CompoundsDocument8 pages(14796813 - Journal of Molecular Endocrinology) Larval Zebrafish As A Model For Glucose Metabolism - Expression of Phosphoenolpyruvate Carboxykinase As A Marker For Exposure To Anti-Diabetic CompoundsGREESTYNo ratings yet
- Skaldin2018 PDFDocument25 pagesSkaldin2018 PDFMaksim SkaldinNo ratings yet
- ISSN 1756-2228 Journal Article on Prostaglandins and the Reproductive CycleDocument36 pagesISSN 1756-2228 Journal Article on Prostaglandins and the Reproductive CycleanakfkubNo ratings yet
- Kumar 1989Document5 pagesKumar 1989Caio OliveiraNo ratings yet
- BMP-2 and CDMP-2: Stimulation of Chondrocyte Production of ProteoglycanDocument8 pagesBMP-2 and CDMP-2: Stimulation of Chondrocyte Production of ProteoglycanFrank ZhouNo ratings yet
- Aerts (2008) CMLS 65, 1933-Ydc1p Ceramidase Triggers Organelle Fragmentation, Apoptosis and Aging in YeastsDocument10 pagesAerts (2008) CMLS 65, 1933-Ydc1p Ceramidase Triggers Organelle Fragmentation, Apoptosis and Aging in YeastsDario FernándezNo ratings yet
- Articulo 1Document16 pagesArticulo 1Juliana CorreaNo ratings yet
- PKD2 eIF2a FASEB J 2013Document12 pagesPKD2 eIF2a FASEB J 2013Xing-Zhen ChenNo ratings yet
- Thermogenic Activity of UCP1 in Human White Fat-Derived Beige Adipocytes 2015 ResearchDocument10 pagesThermogenic Activity of UCP1 in Human White Fat-Derived Beige Adipocytes 2015 Research畏No ratings yet
- Prostaglandin: E - AlprostadilDocument34 pagesProstaglandin: E - AlprostadilJul Hasratman MusaNo ratings yet
- Chen S - 2017Document10 pagesChen S - 2017Tiago OgaitNo ratings yet
- Biochemical and Biophysical Research CommunicationsDocument7 pagesBiochemical and Biophysical Research Communicationsjuan felipeNo ratings yet
- Articulo IiDocument7 pagesArticulo IiOscarDavidGordilloGonzalezNo ratings yet
- Bio Lre Prod 1157Document8 pagesBio Lre Prod 1157Erika Lyz Vanessa Mateo P�rezNo ratings yet
- Hernandez 2012Document4 pagesHernandez 2012Christina MountakiNo ratings yet
- 15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Document7 pages15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Roy GoldenNo ratings yet
- FGF Signaling Pathway and Apert Syndrome MutationsDocument1 pageFGF Signaling Pathway and Apert Syndrome Mutationsamanda liNo ratings yet
- 124 13236FPDocument11 pages124 13236FPFarmasi SMKNegeri1SambiNo ratings yet
- Purification and Properties of Glucose 6-PhosphateDocument7 pagesPurification and Properties of Glucose 6-PhosphateLuceroNo ratings yet
- DiMarzo, Tumoral MacrophagesDocument35 pagesDiMarzo, Tumoral MacrophagesМатиас Себальос ГусманNo ratings yet
- The Relationship Between Alexithymia and Intolerance of Uncertainty in Anorexia NervosaDocument7 pagesThe Relationship Between Alexithymia and Intolerance of Uncertainty in Anorexia NervosaRicardo EscNo ratings yet
- Exploring The Clinical Utility of The DSM-5 Conduct Disorder Specifier of With Limited Prosocial Emotions' in An Adolescent Inpatient SampleDocument16 pagesExploring The Clinical Utility of The DSM-5 Conduct Disorder Specifier of With Limited Prosocial Emotions' in An Adolescent Inpatient SampleRicardo EscNo ratings yet
- Altered Behavior in Mice With Deletion of The Alpha2-Antiplasmin GeneDocument8 pagesAltered Behavior in Mice With Deletion of The Alpha2-Antiplasmin GeneRicardo EscNo ratings yet
- (2015) Marsh - 2015 - Clinical IntuitionsDocument13 pages(2015) Marsh - 2015 - Clinical IntuitionsRicardo EscNo ratings yet
- The Fibrinolytic System: Mysteries and Opportunities: Robert L. Medcalf, Charithani B. KeragalaDocument7 pagesThe Fibrinolytic System: Mysteries and Opportunities: Robert L. Medcalf, Charithani B. KeragalaRicardo EscNo ratings yet
- American University of BeirutDocument54 pagesAmerican University of BeirutRicardo EscNo ratings yet
- Brain-Based Biotypes of Psychiatric Vulnerability in The Acute Aftermath of TraumaDocument13 pagesBrain-Based Biotypes of Psychiatric Vulnerability in The Acute Aftermath of TraumaRicardo EscNo ratings yet
- 494882Document9 pages494882Ricardo EscNo ratings yet
- Journal Pre-Proof: Journal of The American Academy of Child & Adolescent PsychiatryDocument40 pagesJournal Pre-Proof: Journal of The American Academy of Child & Adolescent PsychiatryRicardo EscNo ratings yet
- Psychosocial and Psychological Interventions For RDocument12 pagesPsychosocial and Psychological Interventions For RRicardo EscNo ratings yet
- Psychiatric Leadership in Dispositional Capacity AssessmentsDocument3 pagesPsychiatric Leadership in Dispositional Capacity AssessmentsRicardo EscNo ratings yet
- Adrafinil: Psychostimulant and Purported Nootropic?: Drug ReviewDocument3 pagesAdrafinil: Psychostimulant and Purported Nootropic?: Drug ReviewRicardo EscNo ratings yet
- Monk, Et Al - Prenatal Developmental Origins of Future Psychopathology (2020)Document31 pagesMonk, Et Al - Prenatal Developmental Origins of Future Psychopathology (2020)Ricardo EscNo ratings yet
- Samuel J Millard, Jeremy S Lum, Francesca Fernandez, Katrina Weston-Green and Kelly A NewellDocument14 pagesSamuel J Millard, Jeremy S Lum, Francesca Fernandez, Katrina Weston-Green and Kelly A NewellRicardo EscNo ratings yet
- (2020) Perinatal Mental HealthDocument15 pages(2020) Perinatal Mental HealthRicardo EscNo ratings yet
- Effective Psychological Interventions For RelapseDocument2 pagesEffective Psychological Interventions For RelapseRicardo EscNo ratings yet
- Kovner, Et Al - Cortico-Limbic Interactions Relevant To Psychopathology (2019)Document13 pagesKovner, Et Al - Cortico-Limbic Interactions Relevant To Psychopathology (2019)Ricardo EscNo ratings yet
- What Is Mental Disorder Developing An Embodied Embedded and Enactive PsychopathologyDocument215 pagesWhat Is Mental Disorder Developing An Embodied Embedded and Enactive PsychopathologyRicardo EscNo ratings yet
- Lebowitz & Appelbaum - Biomedical Explanations of Psychopathology (2019)Document26 pagesLebowitz & Appelbaum - Biomedical Explanations of Psychopathology (2019)Ricardo EscNo ratings yet
- Bogdan, Et Al - Polygenic Risk Scores in Clinical Psychology (2018)Document42 pagesBogdan, Et Al - Polygenic Risk Scores in Clinical Psychology (2018)Ricardo EscNo ratings yet
- Common neural circuit disruptions in emotional processing across psychiatric disordersDocument11 pagesCommon neural circuit disruptions in emotional processing across psychiatric disordersRicardo EscNo ratings yet
- Suicidal Ideation - StatPearls - NCBI BookshelfDocument13 pagesSuicidal Ideation - StatPearls - NCBI BookshelfRicardo EscNo ratings yet
- Early Diabetic Risk Prediction Using Machine Learning Classification TechniquesDocument6 pagesEarly Diabetic Risk Prediction Using Machine Learning Classification TechniquesInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- Masterbatch Buyers Guide PDFDocument8 pagesMasterbatch Buyers Guide PDFgurver55No ratings yet
- Barelwiyah, Barelvi Chapter 1 (Part 2 of 5)Document31 pagesBarelwiyah, Barelvi Chapter 1 (Part 2 of 5)Dawah ChannelNo ratings yet
- DLL Grade7 First 1solutions ConcentrationDocument5 pagesDLL Grade7 First 1solutions ConcentrationJaneth de JuanNo ratings yet
- 2nd QUARTER EXAMINATION IN P. E 2019-2020Document3 pages2nd QUARTER EXAMINATION IN P. E 2019-2020Lyzl Mahinay Ejercito MontealtoNo ratings yet
- WEEK 8. - Gen Math-PortfolioDocument12 pagesWEEK 8. - Gen Math-PortfolioOreo ProductionsNo ratings yet
- Corporate Governance in SMEsDocument18 pagesCorporate Governance in SMEsSana DjaanineNo ratings yet
- Organic Facial Remedies Versus Inorganic Facial RemediesDocument13 pagesOrganic Facial Remedies Versus Inorganic Facial Remediesapi-271179911No ratings yet
- Eagle Test ReportDocument25 pagesEagle Test ReportMuhammad FahadNo ratings yet
- State of Education in Tibet 2003Document112 pagesState of Education in Tibet 2003Tibetan Centre for Human Rights and DemocracyNo ratings yet
- Casestudy3 Hbo MaDocument2 pagesCasestudy3 Hbo Ma132345usdfghjNo ratings yet
- Inventory Storage and Retrieval System PatentDocument15 pagesInventory Storage and Retrieval System PatentdevanasokaNo ratings yet
- Inertial Reference Frames: Example 1Document2 pagesInertial Reference Frames: Example 1abhishek murarkaNo ratings yet
- Administracion Una Perspectiva Global Y Empresarial Resumen Por CapitulosDocument7 pagesAdministracion Una Perspectiva Global Y Empresarial Resumen Por Capitulosafmqqaepfaqbah100% (1)
- U-KLEEN Moly Graph MsdsDocument2 pagesU-KLEEN Moly Graph MsdsShivanand MalachapureNo ratings yet
- Manual Handling Risk Assessment ProcedureDocument6 pagesManual Handling Risk Assessment ProcedureSarfraz RandhawaNo ratings yet
- XXCCCDocument17 pagesXXCCCwendra adi pradanaNo ratings yet
- Project Report Software and Web Development Company: WWW - Dparksolutions.inDocument12 pagesProject Report Software and Web Development Company: WWW - Dparksolutions.inRavi Kiran Rajbhure100% (1)
- NSX 100-630 User ManualDocument152 pagesNSX 100-630 User Manualagra04100% (1)
- Building Primitive Traps & SnaresDocument101 pagesBuilding Primitive Traps & SnaresJoseph Madr90% (10)
- Document-SAP EWM For Fashion 1.0: 1.general IntroductionDocument3 pagesDocument-SAP EWM For Fashion 1.0: 1.general IntroductionAnonymous u3PhTjWZRNo ratings yet
- Circuit AnalysisDocument98 pagesCircuit Analysisahtisham shahNo ratings yet
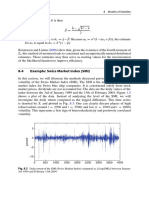
- 8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityDocument3 pages8.4 Example: Swiss Market Index (SMI) : 188 8 Models of VolatilityNickesh ShahNo ratings yet
- Mechanical Engineer with Experience in Heavy Mining Machinery MaintenanceDocument1 pageMechanical Engineer with Experience in Heavy Mining Machinery MaintenanceCertified Rabbits LoverNo ratings yet
- Clarinet Lecture Recital - Jude StefanikDocument35 pagesClarinet Lecture Recital - Jude Stefanikapi-584164068No ratings yet
- 20ME901 Automobile Engineering Unit 3Document74 pages20ME901 Automobile Engineering Unit 36044 sriramNo ratings yet
- Oas Community College: Republic of The Philippines Commission On Higher Education Oas, AlbayDocument22 pagesOas Community College: Republic of The Philippines Commission On Higher Education Oas, AlbayJaycel NepalNo ratings yet
- GVB 1Document8 pagesGVB 1Aaron SylvestreNo ratings yet
- Electrical Design Report For Mixed Use 2B+G+7Hotel BuildingDocument20 pagesElectrical Design Report For Mixed Use 2B+G+7Hotel BuildingEyerusNo ratings yet
- Athenaze 1 Chapter 7a Jun 18th 2145Document3 pagesAthenaze 1 Chapter 7a Jun 18th 2145maverickpussNo ratings yet