You might also like
- Mitosis Is A Process Where A Single Cell Divides Into Two Identical Daughter Cells (Cell Division)Document10 pagesMitosis Is A Process Where A Single Cell Divides Into Two Identical Daughter Cells (Cell Division)Holly Mc Kenzie Layson DalmanNo ratings yet
- Lab Report Bio Experiment 3 - Reproductive, Endocrine & Respiratory System (Group 4)Document15 pagesLab Report Bio Experiment 3 - Reproductive, Endocrine & Respiratory System (Group 4)FARHAH BATRISYIA ABDUL RAHIMNo ratings yet
- Catalogue For TubingDocument1 pageCatalogue For Tubingprabhakaran.cNo ratings yet
- Advance Strategic Marketing: Project Report of Nayatel.Document46 pagesAdvance Strategic Marketing: Project Report of Nayatel.Omer Abbasi60% (15)
- Importance of Human Reproductive SystemDocument3 pagesImportance of Human Reproductive Systemshn mzkNo ratings yet
- Embryology: DR - Kumar Satish Ravi Assist. Prof. & HeadDocument45 pagesEmbryology: DR - Kumar Satish Ravi Assist. Prof. & HeadDr.Kumar Satish RaviNo ratings yet
- Module 3 CytogeneticsDocument23 pagesModule 3 CytogeneticsFrances Riane SimoyNo ratings yet
- The Reproductive SystemDocument68 pagesThe Reproductive SystemSamuelNo ratings yet
- Male & Female Reproductive System: Dr. Zainab Neamat Al-TaeeDocument46 pagesMale & Female Reproductive System: Dr. Zainab Neamat Al-TaeeIn UsNo ratings yet
- Reproductive SystemDocument14 pagesReproductive SystemMichael NyaongoNo ratings yet
- Human Reproduction Unit Review WorksheetDocument25 pagesHuman Reproduction Unit Review Worksheetmofid monirNo ratings yet
- Chapter XII ReproductiveDocument17 pagesChapter XII ReproductiveHannah DiñosoNo ratings yet
- Biology Notes: Key PointsDocument15 pagesBiology Notes: Key PointsAngelineBagtasNo ratings yet
- Lecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyDocument8 pagesLecture Notes Module - 3 - Male Reproductive Anatomy and PhysiologyMariaImranNo ratings yet
- Chapter 19 - ReproductiveDocument18 pagesChapter 19 - Reproductiveapi-220531452No ratings yet
- Physiology II - PresentationDocument11 pagesPhysiology II - PresentationalshmryalmwydNo ratings yet
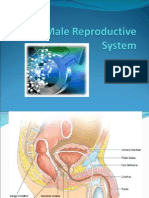
- Male Reproductive SystemDocument49 pagesMale Reproductive SystemRinkish Dalliah50% (2)
- The Male Genital SystemDocument5 pagesThe Male Genital SystemDr-AymanBahgatNo ratings yet
- Embryology: Dr. Caswell HachabizwaDocument67 pagesEmbryology: Dr. Caswell HachabizwaShammar NyirendaNo ratings yet
- MRS LabDocument6 pagesMRS Lab2012000600No ratings yet
- Department of Morphology: Penza State University Medical InstituteDocument18 pagesDepartment of Morphology: Penza State University Medical InstituteunnatiNo ratings yet
- Chapter 28 - The Reproductive SystemDocument7 pagesChapter 28 - The Reproductive Systemtomorrow.today.yesterday .yesterdayNo ratings yet
- Reproductive Systems A. B. C. DDocument6 pagesReproductive Systems A. B. C. DJamie BagundolNo ratings yet
- Human Anatomy,: First Edition Mckinley & O'LoughlinDocument68 pagesHuman Anatomy,: First Edition Mckinley & O'LoughlinnannaraNo ratings yet
- Division of A Parent Cell To Produce To Genetically Identical Daughter CellsDocument6 pagesDivision of A Parent Cell To Produce To Genetically Identical Daughter Cellseurovision fanNo ratings yet
- Neiron GametogenesisDocument6 pagesNeiron GametogenesisJose MartinezNo ratings yet
- Chapter 25 - Reproduction and Human DevelopmentDocument33 pagesChapter 25 - Reproduction and Human DevelopmentmaleskunNo ratings yet
- Week 16 - Reproductive SystemDocument71 pagesWeek 16 - Reproductive SystemGeevee Naganag VentulaNo ratings yet
- Anatomy and Physiology of Male Reproductive SystemDocument6 pagesAnatomy and Physiology of Male Reproductive SystemZander Ragasa GuzmanNo ratings yet
- Reproduction in AnimalsDocument12 pagesReproduction in AnimalsYuh moddaNo ratings yet
- SpermatogenesisDocument31 pagesSpermatogenesiselzenyNo ratings yet
- 11 MaleDocument14 pages11 MaleJasmine KaurNo ratings yet
- The Reproductive Systems AnatomyDocument3 pagesThe Reproductive Systems AnatomyLilian JerotichNo ratings yet
- The Reproductive SystemDocument15 pagesThe Reproductive SystemMostafa Galal El Din100% (1)
- Human Reproduction and Reproductive HealthDocument16 pagesHuman Reproduction and Reproductive HealthHemanth GowdaNo ratings yet
- Gametogenesis, Gamete Transport & FertilizationDocument56 pagesGametogenesis, Gamete Transport & FertilizationvictorNo ratings yet
- Intoduction To Embryology, Testis & Spermatogenesis - Nto, Nto JohnsonDocument26 pagesIntoduction To Embryology, Testis & Spermatogenesis - Nto, Nto JohnsonigboegwummesomaNo ratings yet
- Group 5 Gametes and FertilizationDocument74 pagesGroup 5 Gametes and FertilizationLira PagaraNo ratings yet
- Male Reproductive OrganDocument43 pagesMale Reproductive OrganIsrahia CopiocoNo ratings yet
- The Reproductive OrgansDocument39 pagesThe Reproductive OrgansjoaNo ratings yet
- Human Life Cycle & Male Reproductive System: Dr. Muhjah Falah Hasan PH.D in Infertility & Clinical ReproductionDocument32 pagesHuman Life Cycle & Male Reproductive System: Dr. Muhjah Falah Hasan PH.D in Infertility & Clinical Reproductionsuad aliNo ratings yet
- Lab Report Bio Experiment 5 (Mouse Dissection)Document14 pagesLab Report Bio Experiment 5 (Mouse Dissection)Hakim SunaeNo ratings yet
- Spermatogenesis 1Document28 pagesSpermatogenesis 1LellllNo ratings yet
- Science 10 3rd Grading Module 1 REPRODUCTIVESYSTEMDocument32 pagesScience 10 3rd Grading Module 1 REPRODUCTIVESYSTEMvinesse100% (1)
- Anatomy and Physiology of Male Reproductive SystemDocument8 pagesAnatomy and Physiology of Male Reproductive SystemAdor AbuanNo ratings yet
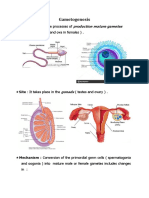
- GametogenesisDocument35 pagesGametogenesisJason TulipatNo ratings yet
- Anatomy of The Male Reproductive SystemDocument17 pagesAnatomy of The Male Reproductive SystemFarhan haiderNo ratings yet
- 1 GametogenesisDocument41 pages1 Gametogenesishaladaqqa1No ratings yet
- Gametogenesis Adalah Proses Pembentukan Gamet Atau Sel Kelamin. Sel GametDocument12 pagesGametogenesis Adalah Proses Pembentukan Gamet Atau Sel Kelamin. Sel Gametadelina savitriNo ratings yet
- SpermatogenesisDocument13 pagesSpermatogenesiszuha khanNo ratings yet
- Spermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyDocument41 pagesSpermatogenesis: Feriha Ercan, Ph.D. Dept. Histology and EmbryologyErdem AltunNo ratings yet
- Plant: Reproductive SystemDocument3 pagesPlant: Reproductive Systemchan_1016No ratings yet
- Spermatogenesis WPS Office 1Document12 pagesSpermatogenesis WPS Office 1nb6pd5rvnxNo ratings yet
- KM GametogenesisDocument42 pagesKM GametogenesisKALAMYA BRIAN BWAYONo ratings yet
- Histology of The Male Reproductive SystemDocument18 pagesHistology of The Male Reproductive SystemMuhammad JameelNo ratings yet
- EmbryologyDocument89 pagesEmbryologyPythias MumaNo ratings yet
- Reproduction and DevelopmentDocument121 pagesReproduction and DevelopmentAmethystNo ratings yet
- GametogenesisDocument5 pagesGametogenesisjabreen ahmadNo ratings yet
- Formation of GametesDocument27 pagesFormation of GametesKalo kajiNo ratings yet
- Reproductive System Powerpoint LectureDocument55 pagesReproductive System Powerpoint LectureJames DaurayNo ratings yet
- General Embryology 2022Document130 pagesGeneral Embryology 2022KAMAL ALSOFIANYNo ratings yet
- HIS - Topic 1-2 - PorzaDocument10 pagesHIS - Topic 1-2 - PorzaZuhri PorzaNo ratings yet
- Happ PrelimsDocument12 pagesHapp PrelimsZuhri PorzaNo ratings yet
- Heart: StructureDocument11 pagesHeart: StructureKert trocioNo ratings yet
- HAPP Supplementary Notes - Urinary SystemDocument15 pagesHAPP Supplementary Notes - Urinary SystemZuhri PorzaNo ratings yet
- HAPP Supplementary Notes - BloodDocument14 pagesHAPP Supplementary Notes - BloodZuhri PorzaNo ratings yet
- HAPP Supplementary Notes - Lymphatic SystemDocument14 pagesHAPP Supplementary Notes - Lymphatic SystemZuhri PorzaNo ratings yet
- GSKDocument22 pagesGSKChaudhary Hassan ArainNo ratings yet
- Seven Elements of Effective NegotiationsDocument3 pagesSeven Elements of Effective NegotiationsKilik GantitNo ratings yet
- Microtronics Technologies: GSM Based Vehicle Theft Detection SystemDocument3 pagesMicrotronics Technologies: GSM Based Vehicle Theft Detection Systemابراهيم الثوبريNo ratings yet
- II If en April2015 EquitycompoundersDocument8 pagesII If en April2015 EquitycompoundersbgyggghjkkNo ratings yet
- Jasa Marga TBK - Billingual - 31 - Des - 2018 - JSMR PDFDocument227 pagesJasa Marga TBK - Billingual - 31 - Des - 2018 - JSMR PDFSjamsulHarunNo ratings yet
- Cancer Registry Standard Operating ProceduresDocument3 pagesCancer Registry Standard Operating ProceduresAnan AghbarNo ratings yet
- AX Performance Tools1Document21 pagesAX Performance Tools1Htun Lin AungNo ratings yet
- COT RPMS Rating Sheet For T I III For SY 2021 2022 JhanzDocument2 pagesCOT RPMS Rating Sheet For T I III For SY 2021 2022 Jhanzjhancelle golosindaNo ratings yet
- Survey-Questionnaire For The Study "Factors Affecting The Level of Comprehension in English of Grade 3 Students School Year 2021-2023"Document13 pagesSurvey-Questionnaire For The Study "Factors Affecting The Level of Comprehension in English of Grade 3 Students School Year 2021-2023"Rosalinda SamongNo ratings yet
- Afternoon Quiz Set 4Document66 pagesAfternoon Quiz Set 4pchakkrapani100% (1)
- Bba-Mq Tias 2023-24Document2 pagesBba-Mq Tias 2023-24Chris PresleyNo ratings yet
- Dialer AdminDocument5 pagesDialer AdminNaveenNo ratings yet
- TDBFP - Gear Pump API 676 PDFDocument42 pagesTDBFP - Gear Pump API 676 PDFRamon A. Ruiz O.No ratings yet
- (Separation Pay - Legal Termination Due To Authorized Causes) National Federation of Labor v. Court of Appeals, 440 SCRA 604 (2004Document6 pages(Separation Pay - Legal Termination Due To Authorized Causes) National Federation of Labor v. Court of Appeals, 440 SCRA 604 (2004Jude FanilaNo ratings yet
- NEBULIZATIONDocument2 pagesNEBULIZATIONMae ValenzuelaNo ratings yet
- 4 Komponen SDSSDocument9 pages4 Komponen SDSSRebecha Parsellya100% (1)
- Eligibility Conditions: Advertisement For Regular Commission in Pakistan Army Through 136 Pma Long CourseDocument5 pagesEligibility Conditions: Advertisement For Regular Commission in Pakistan Army Through 136 Pma Long CourseHusnain IshtiaqNo ratings yet
- Name and Logo Design Contest For Public Wi-Fi Network Services Terms & ConditionsDocument2 pagesName and Logo Design Contest For Public Wi-Fi Network Services Terms & ConditionsAc RaviNo ratings yet
- Making Friends British English StudentDocument8 pagesMaking Friends British English StudentMily ArrayaNo ratings yet
- Assembly Lab 1Document13 pagesAssembly Lab 1Natpro CoolNo ratings yet
- Government and BusinessDocument2 pagesGovernment and BusinessJoshua BrownNo ratings yet
- XDocument2 pagesXSophiaFrancescaEspinosaNo ratings yet
- Model Teaching CompetenciesDocument12 pagesModel Teaching CompetenciesTeachers Without BordersNo ratings yet
- BOLBOK (1st)Document10 pagesBOLBOK (1st)Mj EndozoNo ratings yet
- OfficeServ 7200 Service Manual - Ed.0Document351 pagesOfficeServ 7200 Service Manual - Ed.0Richard WybornNo ratings yet
- DBMS NotesDocument6 pagesDBMS Notesᴠɪᴄɪᴏᴜs ᴄᴜʀsᴇNo ratings yet
- Name - ESM 2104 Sample Final ExamDocument7 pagesName - ESM 2104 Sample Final ExamCOCO12432345No ratings yet
- Mcmurdo S4 SART User GuideDocument20 pagesMcmurdo S4 SART User Guidesathish kumar100% (2)