You might also like
- Systematic Osteology 2020Document20 pagesSystematic Osteology 2020KalangiIrushikaNo ratings yet
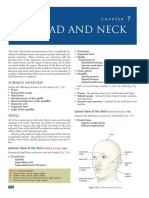
- Chapter07 TheHead&Neck 150 165 PDFDocument16 pagesChapter07 TheHead&Neck 150 165 PDFPopa AlexandraNo ratings yet
- All Bony LandmarkDocument34 pagesAll Bony Landmarkvickytang22072002No ratings yet
- Anatomy of the SkullDocument29 pagesAnatomy of the Skullaya abdullahNo ratings yet
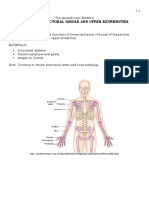
- Lab 7 Pectoral Girdle 2Document4 pagesLab 7 Pectoral Girdle 2Bechris BambiNo ratings yet
- Integrative Functions of The Nervous SystemDocument18 pagesIntegrative Functions of The Nervous Systempuchio100% (1)
- Lec.2 H&N Anatomy, BuraqDocument55 pagesLec.2 H&N Anatomy, BuraqAkramNo ratings yet
- Anatomy and Physiology 6th Edition Marieb Solutions ManualDocument20 pagesAnatomy and Physiology 6th Edition Marieb Solutions Manualstephenthanh1huo100% (28)
- Anatomy and Physiology 6th Edition Marieb Solutions Manual Full Chapter PDFDocument41 pagesAnatomy and Physiology 6th Edition Marieb Solutions Manual Full Chapter PDFserenafinnodx100% (9)
- Anatomy Lect of Skull 2Document36 pagesAnatomy Lect of Skull 2sanullah123khan.13No ratings yet
- 1anatomy of The Nervous System Part I PDFDocument79 pages1anatomy of The Nervous System Part I PDFOmar AlemánNo ratings yet
- Virtual Laboratory Activity Worksheet On Central Nervous SystemDocument6 pagesVirtual Laboratory Activity Worksheet On Central Nervous SystemAngela ParaisoNo ratings yet
- l15 CerebrumDocument31 pagesl15 CerebrumDr. Pinki RaiNo ratings yet
- Johns 2014Document21 pagesJohns 2014SAYDE ILEANA NAJERA DOMINGUEZNo ratings yet
- NEUROANATOMYDocument34 pagesNEUROANATOMYRavanno Fanizza HarahapNo ratings yet
- Yyyyycrash Course Anatomy - Head & NeckDocument52 pagesYyyyycrash Course Anatomy - Head & Neck黃芳昌No ratings yet
- 03 Meninges, Neuron, NMJ, CSF (Students Copy)Document2 pages03 Meninges, Neuron, NMJ, CSF (Students Copy)Hillary DangananNo ratings yet
- Color Atlas of the BrainDocument8 pagesColor Atlas of the BrainMWNo ratings yet
- Frontal LobeDocument39 pagesFrontal LobeVIKAS PUNIANo ratings yet
- Open AnatomyDocument50 pagesOpen AnatomyalukiasuNo ratings yet
- Ilovepdf MergedDocument65 pagesIlovepdf Mergedrogegir407No ratings yet
- 17 Skull Mandible Vertebrae DR - GosaiDocument69 pages17 Skull Mandible Vertebrae DR - GosaiDr.B.B.GosaiNo ratings yet
- Deiuliis2019 VERTEBRADOS LA PERCA 4Document10 pagesDeiuliis2019 VERTEBRADOS LA PERCA 4Fernando Melo GarciaNo ratings yet
- L15 CerebrumDocument27 pagesL15 CerebrumEvilneko1No ratings yet
- Mini AtlasDocument28 pagesMini AtlasAlex WoodwardNo ratings yet
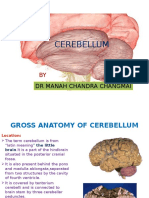
- Cerebellum: DR Manah Chandra ChangmaiDocument60 pagesCerebellum: DR Manah Chandra Changmainairda12No ratings yet
- Elbow Complex (Kinesiology)Document2 pagesElbow Complex (Kinesiology)Kimmybee Garcia50% (2)
- Nerve - H&N PracticeDocument7 pagesNerve - H&N PracticeThục MỹNo ratings yet
- 02 SkullDocument19 pages02 Skullapi-19641337No ratings yet
- Chapter 8 Lab Assignment - Appendicular Skeleton LabelingDocument6 pagesChapter 8 Lab Assignment - Appendicular Skeleton Labelingadriana blanco galianoNo ratings yet
- Anatomia para Estética Parte 1Document45 pagesAnatomia para Estética Parte 1Rodolpho VilelaNo ratings yet
- Materials Needed:: Nervous System Laboratory ProcedureDocument7 pagesMaterials Needed:: Nervous System Laboratory ProcedureRegina GambayanNo ratings yet
- Chapter 5Document41 pagesChapter 5Reneilwe MoshidiNo ratings yet
- 1 - The SkullDocument68 pages1 - The Skullaboody omerNo ratings yet
- p6. The Cerebral CortexDocument99 pagesp6. The Cerebral CortexGedefaw TegegneNo ratings yet
- Lab+Exam+1+Worksheet+ +terminology 1Document11 pagesLab+Exam+1+Worksheet+ +terminology 1jharvillgreeneNo ratings yet
- Cerebral Cortex1Document37 pagesCerebral Cortex1xwrmaliaramNo ratings yet
- (849-853) Component PartsDocument5 pages(849-853) Component PartsMỉm CườiNo ratings yet
- Poster01 Skeletal-System-Anterior Ledger11x17Document1 pagePoster01 Skeletal-System-Anterior Ledger11x17jeffrin alexanderNo ratings yet
- Gross Structure of The BrainDocument60 pagesGross Structure of The BrainPurnomo Ponco NugrohoNo ratings yet
- Anatomy of The Human BodyDocument6 pagesAnatomy of The Human BodyYara AmadeNo ratings yet
- Scalp (2) PP - 240218 - 203325Document16 pagesScalp (2) PP - 240218 - 203325Hager AbouzaidNo ratings yet
- 3 Face and Parotid SlidesDocument23 pages3 Face and Parotid SlidesAmbg GhalyNo ratings yet
- معمل upper limb د علي البغدادي الجزء الاولDocument8 pagesمعمل upper limb د علي البغدادي الجزء الاولAbdulaziz MohammedNo ratings yet
- Osteology Summary NotesDocument25 pagesOsteology Summary Notesmilex75862No ratings yet
- Bones & Joints of Head & NeckDocument91 pagesBones & Joints of Head & NeckSalman KhanNo ratings yet
- Lecture 12 Osteology Upper LimbDocument25 pagesLecture 12 Osteology Upper LimbaidfajdfiajNo ratings yet
- Lab 4 - ForearmDocument44 pagesLab 4 - ForearmTayai LengimaNo ratings yet
- The Axial SkeletonDocument57 pagesThe Axial SkeletonJudithNo ratings yet
- Cerebral Cortex: Medical Neuroscience Dr. WiegandDocument24 pagesCerebral Cortex: Medical Neuroscience Dr. WiegandUzma KhanNo ratings yet
- CEPHALOMETRICS: A GUIDE TO CEPHALOMETRIC ANALYSISDocument141 pagesCEPHALOMETRICS: A GUIDE TO CEPHALOMETRIC ANALYSISshyama pramodNo ratings yet
- Laboratory Manual For Anatomy and Physiology 6Th Edition Marieb Solutions Manual Full Chapter PDFDocument28 pagesLaboratory Manual For Anatomy and Physiology 6Th Edition Marieb Solutions Manual Full Chapter PDFJohnathanFitzgeraldnwoa100% (10)
- Laboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions ManualDocument7 pagesLaboratory Manual For Anatomy and Physiology 6th Edition Marieb Solutions Manualhanhgiaoghhr100% (33)
- Varma 1st Exam NotesDocument7 pagesVarma 1st Exam NotesKarthick KanagarajNo ratings yet
- LimffffffffDocument77 pagesLimffffffffНемосјановић ЋудмилаNo ratings yet
- Prevention & Management of Vision Loss Relating To Facial Filler InjectionsDocument6 pagesPrevention & Management of Vision Loss Relating To Facial Filler InjectionsDenis CartottoNo ratings yet
- Anatomi HidungDocument28 pagesAnatomi HidungferyantoachmadNo ratings yet
- Material (BBC)Document43 pagesMaterial (BBC)vaniaNo ratings yet
- Anatomi System ScletalDocument32 pagesAnatomi System ScletalAndre HawkNo ratings yet
- Structure and Function of Declarative and Nondeclarative Memory SystemsDocument8 pagesStructure and Function of Declarative and Nondeclarative Memory Systemscecilia martinezNo ratings yet
- Calculation Sheet: For Construction Project of Villa Consists of Ground, Frist Floor and RoofDocument62 pagesCalculation Sheet: For Construction Project of Villa Consists of Ground, Frist Floor and Roofقيس عبدالرحمن قيسNo ratings yet
- Aplikasi Kapur Pertanian Untuk Peningkatan Produksi Tanaman Padi Di Tanah Sawah AluvialDocument7 pagesAplikasi Kapur Pertanian Untuk Peningkatan Produksi Tanaman Padi Di Tanah Sawah AluvialChesi HardianaNo ratings yet
- The Limbic Brain. Andrew Lautin PDFDocument152 pagesThe Limbic Brain. Andrew Lautin PDFAntonio D LazodelaVegaNo ratings yet
- Clinical Neuropsychology (Kessels, Roy Eling, Paul Ponds Etc.) (Z-Lib - Org) - 1Document625 pagesClinical Neuropsychology (Kessels, Roy Eling, Paul Ponds Etc.) (Z-Lib - Org) - 1Michelle van DienstNo ratings yet
- Concrete Mix Design - STFA - 09.06.2009Document40 pagesConcrete Mix Design - STFA - 09.06.2009masahinNo ratings yet
- PTSD Prefrontal Cortex and AmigdalaDocument13 pagesPTSD Prefrontal Cortex and AmigdalaMeryem FilizNo ratings yet
- Nature Neuroscience December 2009Document149 pagesNature Neuroscience December 2009patricia_666No ratings yet
- Physiology of Memory and LearningDocument48 pagesPhysiology of Memory and LearningWahyoe PoeshNo ratings yet
- (Developmental Neuroscience) Anthony T. Campagnoni-Brain Development With A Focus On The Cortex - Developmental Neuroscience Vol 25 Issue 2-4-S Karger Pub (2003) PDFDocument215 pages(Developmental Neuroscience) Anthony T. Campagnoni-Brain Development With A Focus On The Cortex - Developmental Neuroscience Vol 25 Issue 2-4-S Karger Pub (2003) PDFFrancisco DiezNo ratings yet
- Cognitive Assessment For Clinicians 3rd EditionDocument298 pagesCognitive Assessment For Clinicians 3rd EditionInes Strenja Linić100% (3)
- Stress and HealthDocument40 pagesStress and HealthSomrudee SaiyudthongNo ratings yet
- DM Neurology ThesisDocument7 pagesDM Neurology ThesisFindSomeoneToWriteMyPaperLowell100% (2)
- Major Brain Regions and Their Major Interconnections (김Document2 pagesMajor Brain Regions and Their Major Interconnections (김신윤상0% (1)
- Bliss and Lomo (1973)Document26 pagesBliss and Lomo (1973)delahunty123No ratings yet
- Hebbian Plasticity: Ã 2009 Elsevier Ltd. All Rights ReservedDocument8 pagesHebbian Plasticity: Ã 2009 Elsevier Ltd. All Rights ReservedCecilia PoganNo ratings yet
- Specialized Astrocytes Mediate Glutamatergic Gliotransmission in The CNSDocument47 pagesSpecialized Astrocytes Mediate Glutamatergic Gliotransmission in The CNSCARDIO 2019No ratings yet
- Anatomy and Function of The HypothalamusDocument13 pagesAnatomy and Function of The HypothalamusNrs Sani Sule MashiNo ratings yet
- Fixed Stage Upright MicroscopeDocument14 pagesFixed Stage Upright MicroscopeAmy AdamsNo ratings yet
- BE Ist Year Sem 1 and 2 - Full Scheme With Syllabus & SU CodeDocument86 pagesBE Ist Year Sem 1 and 2 - Full Scheme With Syllabus & SU CodePÀRMAR parthNo ratings yet
- Kalpa in The PhysiologyDocument58 pagesKalpa in The PhysiologysaidevoNo ratings yet
- Schema Ketronx1Document12 pagesSchema Ketronx1Manrolla89% (9)
- Paxinos The Rat Brain 73Document1 pagePaxinos The Rat Brain 73joseuri2011No ratings yet
- Practical Computer Application Formatting and CalculationsDocument22 pagesPractical Computer Application Formatting and CalculationsBabu Bihari LalNo ratings yet
- 14 GMD - Cycloconverter Control TerminalDocument15 pages14 GMD - Cycloconverter Control TerminalIrving Dirzo Carrillo100% (1)
- Clinical: Dialogu S eDocument144 pagesClinical: Dialogu S eDiego A. Cuadros Torres100% (3)
- Probability CalculatorDocument104 pagesProbability CalculatorNorlly Mohd IsaNo ratings yet
- Alekls 1202Document113 pagesAlekls 1202Joseph SikatumbaNo ratings yet
- Resveratrol Therapy For EpilepsyDocument37 pagesResveratrol Therapy For EpilepsyAlejandro Nico Rondón OrtizNo ratings yet
- George K. Aghajanian and Elaine Sanders-Bush - SerotoninDocument20 pagesGeorge K. Aghajanian and Elaine Sanders-Bush - SerotoninSour60No ratings yet
- The Age of Magical Overthinking: Notes on Modern IrrationalityFrom EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityRating: 4 out of 5 stars4/5 (20)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedRating: 5 out of 5 stars5/5 (78)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (5)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsFrom EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNo ratings yet
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionFrom EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionRating: 4 out of 5 stars4/5 (403)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisFrom EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (42)
- 12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosFrom Everand12 Rules for Life by Jordan B. Peterson - Book Summary: An Antidote to ChaosRating: 4.5 out of 5 stars4.5/5 (207)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsFrom EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsRating: 5 out of 5 stars5/5 (1)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeFrom EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeNo ratings yet
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsFrom EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsRating: 3.5 out of 5 stars3.5/5 (3)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (169)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaFrom EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaRating: 4.5 out of 5 stars4.5/5 (266)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryFrom EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryRating: 4 out of 5 stars4/5 (44)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessFrom EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessRating: 4.5 out of 5 stars4.5/5 (328)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.From EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Rating: 4.5 out of 5 stars4.5/5 (110)
- The Happiness Trap: How to Stop Struggling and Start LivingFrom EverandThe Happiness Trap: How to Stop Struggling and Start LivingRating: 4 out of 5 stars4/5 (1)
- Algorithms to Live By: The Computer Science of Human DecisionsFrom EverandAlgorithms to Live By: The Computer Science of Human DecisionsRating: 4.5 out of 5 stars4.5/5 (722)
- Summary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedFrom EverandSummary: How to Be an Adult in Relationships: The Five Keys to Mindful Loving by David Richo: Key Takeaways, Summary & Analysis IncludedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Secure Love: Create a Relationship That Lasts a LifetimeFrom EverandSecure Love: Create a Relationship That Lasts a LifetimeRating: 5 out of 5 stars5/5 (18)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)