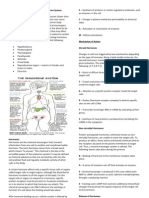
You might also like
- IP-Worksheet-3-Endocrine System The Hypothalamic-Pituitary Axis (Estomagulang)Document2 pagesIP-Worksheet-3-Endocrine System The Hypothalamic-Pituitary Axis (Estomagulang)Filmae Estomagulang0% (1)
- EndoquestionssolvedDocument14 pagesEndoquestionssolvedPuteri Nursyamimi Mohd SabilanNo ratings yet
- Endocrine System MCQDocument19 pagesEndocrine System MCQBijay Kumar MahatoNo ratings yet
- The hypothalamus and pituitary glandDocument13 pagesThe hypothalamus and pituitary glandKiprono Keitany TimothyNo ratings yet
- Pcol Lec Book FinalsDocument61 pagesPcol Lec Book FinalsMarco Sta AnaNo ratings yet
- Biochemistry of Hormones (Part 1)Document31 pagesBiochemistry of Hormones (Part 1)arun231187No ratings yet
- Clinical BiochemistryDocument71 pagesClinical BiochemistryMuhamed ArsalanNo ratings yet
- Phys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESDocument7 pagesPhys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESsine summsun PradhanNo ratings yet
- Chapter - 17 PhysiologyDocument49 pagesChapter - 17 PhysiologyAhsanullah PathanNo ratings yet
- Under The Guidance Of:: Dr. Sandeep Tandon Professor and Head of Dept. of PedodonticsDocument79 pagesUnder The Guidance Of:: Dr. Sandeep Tandon Professor and Head of Dept. of PedodonticsMarivic DianoNo ratings yet
- Dermatologic manifestations of hypopituitarismDocument10 pagesDermatologic manifestations of hypopituitarismWPRN RanasingheNo ratings yet
- The Endocrine System: Hormones, Receptors and RegulationDocument47 pagesThe Endocrine System: Hormones, Receptors and RegulationRifi OktasiaNo ratings yet
- Disorders of The Anterior Pituitary and HypothalamusDocument121 pagesDisorders of The Anterior Pituitary and Hypothalamusbiniam MesfinNo ratings yet
- Endocrine SystemDocument70 pagesEndocrine SystemKELVINNo ratings yet
- Endocrine ResonanceDocument51 pagesEndocrine ResonanceEkta ManglaniNo ratings yet
- Endocrine System: DR - Entissar Mansour 2O16-2017Document162 pagesEndocrine System: DR - Entissar Mansour 2O16-2017Zainab Alshamary100% (1)
- Endocrine System - Part 1 (Robbins)Document28 pagesEndocrine System - Part 1 (Robbins)sarguss14100% (2)
- English Sistem EndocrineDocument9 pagesEnglish Sistem EndocrineNurlaili YaniNo ratings yet
- Cells 10 02664Document13 pagesCells 10 02664Manaloe90No ratings yet
- Hormonal Management of Male InfertilityDocument22 pagesHormonal Management of Male Infertilitydranibalurologoencancun.mxNo ratings yet
- Hypothalamus - Anterior Pituitary and Their HormonesDocument138 pagesHypothalamus - Anterior Pituitary and Their HormonesML Rodriguez100% (1)
- Pituitary Gland Hormones & Their FunctionsDocument35 pagesPituitary Gland Hormones & Their Functionsbhagirath kansara100% (1)
- Endocrine System: EndocrinologyDocument10 pagesEndocrine System: EndocrinologyAnya IgnacioNo ratings yet
- Male ReproductiveDocument10 pagesMale ReproductivevivekNo ratings yet
- Cap.1 - Hypothalamic-Pituitary Sistem 2018 - 2019Document23 pagesCap.1 - Hypothalamic-Pituitary Sistem 2018 - 2019Razvan GeorgeNo ratings yet
- Endocrinology Notes: Veterinary PhysiologyDocument16 pagesEndocrinology Notes: Veterinary PhysiologyBrian AllanNo ratings yet
- Pituitary Blok Endokrin 2019Document40 pagesPituitary Blok Endokrin 2019ratihNo ratings yet
- Pituitary Hormones and Their FunctionsDocument8 pagesPituitary Hormones and Their FunctionsNoreen B. BañagadoNo ratings yet
- EndocrinologyDocument16 pagesEndocrinologyJack Nicholas BusaNo ratings yet
- 4.0 The Pituitary GlandDocument68 pages4.0 The Pituitary GlandHomeground entertainmentNo ratings yet
- Reproductive Physiology PDFDocument70 pagesReproductive Physiology PDFDrbee10No ratings yet
- Functional Anatomy of The Hypothalamicpituitarygonadal Axis andDocument10 pagesFunctional Anatomy of The Hypothalamicpituitarygonadal Axis andEvaNo ratings yet
- Endocrinology Apllied ScienceDocument44 pagesEndocrinology Apllied ScienceAdam PrabowoNo ratings yet
- Endocrinal PathologyDocument81 pagesEndocrinal PathologyshehlaNo ratings yet
- Topic 1 Endocrine SystemDocument140 pagesTopic 1 Endocrine SystemmasdfgNo ratings yet
- Endocrinology PDFDocument11 pagesEndocrinology PDFLawrence Genelago GamboaNo ratings yet
- NeuroendocrinologyDocument58 pagesNeuroendocrinologyyuyu tuptupNo ratings yet
- Endocrine GlandDocument4 pagesEndocrine Glandakash_zizouNo ratings yet
- Anatomy and Physiology-Endocrine SystemDocument5 pagesAnatomy and Physiology-Endocrine SystemEixid Enna YeLikNo ratings yet
- Reproductive MedicineDocument330 pagesReproductive MedicineunknownxemNo ratings yet
- Reproductive Endocrinology FinalDocument98 pagesReproductive Endocrinology FinalChino Paolo SamsonNo ratings yet
- Endokrin MW DitransletDocument7 pagesEndokrin MW DitransletSanta SaintNo ratings yet
- Clinical Chemistry - PITUITARY HORMONES TRANSDocument7 pagesClinical Chemistry - PITUITARY HORMONES TRANSCamella Beatrice Lujan ValleNo ratings yet
- Clinton Hackney Anatomy & Physiology II BctcsDocument65 pagesClinton Hackney Anatomy & Physiology II BctcsJack HolthouseNo ratings yet
- EndokrinDocument70 pagesEndokrinNur Hadi PrayitnoNo ratings yet
- Chapter 34 - Introduction to the Endocrine SystemDocument3 pagesChapter 34 - Introduction to the Endocrine Systemkdayeon018No ratings yet
- UNIT II Endocrine & Metabolic DisordersDocument19 pagesUNIT II Endocrine & Metabolic DisordersAmmar BhattiNo ratings yet
- Drugs Affecting The Endocrine SystemDocument43 pagesDrugs Affecting The Endocrine SystemJustine Vens G. AgustinNo ratings yet
- Pituitary GlandDocument3 pagesPituitary GlandAllyza Marie PeñaNo ratings yet
- Johanna Jane H. Macasero - Endocrine System TranscriptDocument13 pagesJohanna Jane H. Macasero - Endocrine System TranscriptJohanna MacaseroNo ratings yet
- Introduction to EndocrinologyDocument19 pagesIntroduction to EndocrinologyVDiesel99No ratings yet
- Unit2-Hypothalamus and PituitaryDocument80 pagesUnit2-Hypothalamus and Pituitarypranutan739No ratings yet
- European Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyDocument6 pagesEuropean Journal of Endocrinology) Corticotropin-Releasing Hormone PhysiologyBenedictus Yudha BaskaraNo ratings yet
- The Endocrine System: Properties: Mechanisms of Hormone ActionDocument5 pagesThe Endocrine System: Properties: Mechanisms of Hormone ActionaudreyNo ratings yet
- Care of Clients With Problem On EndocrineDocument48 pagesCare of Clients With Problem On EndocrineAngel CauilanNo ratings yet
- The Endocrine System Consists of Hormones Secreted by Glands Traveling Through The Blood To Target TissuesDocument16 pagesThe Endocrine System Consists of Hormones Secreted by Glands Traveling Through The Blood To Target TissuesAljon AniesNo ratings yet
- Endocrine System (Ch7)Document15 pagesEndocrine System (Ch7)moepiNo ratings yet
- Review of Endocrine SystemDocument32 pagesReview of Endocrine SystemJames FNo ratings yet
- An Introduction To EndocrinologyDocument47 pagesAn Introduction To EndocrinologyAmeng GosimNo ratings yet
- Endocrine GlandsDocument36 pagesEndocrine GlandsnasyitahnorzlanNo ratings yet
- EN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)Document24 pagesEN17 - Tumors of The Sellar Region - Acromegaly, Prolactinoma, Gonadotropinoma, Nonfunctioning Pituitary Adenoma, Craniopharyngioma)sbobine.imsNo ratings yet
- Neurochemical Aspects of Hypothalamic FunctionFrom EverandNeurochemical Aspects of Hypothalamic FunctionL MartiniNo ratings yet
- Lecture 2 - Pituitary Hormones and Their Control by The HypothalamusDocument39 pagesLecture 2 - Pituitary Hormones and Their Control by The HypothalamusHomeground entertainmentNo ratings yet
- Pituitary Disorders Throughout The Life Cycle: A Case-Based Guide Susan L. Samson Adriana G. IoachimescuDocument399 pagesPituitary Disorders Throughout The Life Cycle: A Case-Based Guide Susan L. Samson Adriana G. IoachimescuyusranurparlakNo ratings yet
- HGH Effects Growth DevelopmentDocument2 pagesHGH Effects Growth DevelopmentMichael BriscoeNo ratings yet
- Kraemer 2010Document11 pagesKraemer 2010Vukašin Schooly StojanovićNo ratings yet
- Bodybuilding Nutrition 4 - Growth Hormone BoostersDocument10 pagesBodybuilding Nutrition 4 - Growth Hormone BoostersccciiipppNo ratings yet
- Growth Hormone - BDSDocument24 pagesGrowth Hormone - BDSxmarty.fazil100% (1)
- PeptidesDocument19 pagesPeptidesNate HardingNo ratings yet
- Ultimate Peptide GuideDocument23 pagesUltimate Peptide GuideMarzena KarmecckaNo ratings yet
- BCP 14265Document10 pagesBCP 14265Noemi Marino FloresNo ratings yet
- Growth HormoneDocument39 pagesGrowth Hormonesalma100% (1)
- Hormonal Regulation of MetabolismDocument80 pagesHormonal Regulation of MetabolismVivek ChaudharyNo ratings yet
- Introduction to EndocrinologyDocument19 pagesIntroduction to EndocrinologyVDiesel99No ratings yet
- Doi PMC Pmid: Jump Up ToDocument3 pagesDoi PMC Pmid: Jump Up ToKunj TechnicalNo ratings yet
- What Are HormonesDocument16 pagesWhat Are HormonesHarry RoyNo ratings yet
- NASC 4 Fact or BluffDocument33 pagesNASC 4 Fact or BluffJona MangabanNo ratings yet
- Growth Hormone Therapy: Guided By: Dr. Arif VohraDocument45 pagesGrowth Hormone Therapy: Guided By: Dr. Arif VohraMitulNo ratings yet
- Pituitary GlandDocument15 pagesPituitary GlandPutri Agustin Mere100% (1)
- Vety Endocrine PharmaDocument59 pagesVety Endocrine PharmaSunilNo ratings yet
- Endocrine SystemDocument7 pagesEndocrine SystemSolita PorteriaNo ratings yet
- Endocrine System Lecture SummaryDocument193 pagesEndocrine System Lecture SummaryLanaSNo ratings yet
- Pediatric Endocrinology A Practical Clinical Guide, 2E 2013 (PDF) (DR - Carson) VRG PDFDocument615 pagesPediatric Endocrinology A Practical Clinical Guide, 2E 2013 (PDF) (DR - Carson) VRG PDFIonela RobertaNo ratings yet
- Growth Hormone Deficiency - Springer (2016)Document232 pagesGrowth Hormone Deficiency - Springer (2016)Atp FranklinNo ratings yet
- Endocrine System: Clarence L. Nuval, RMT, MDDocument66 pagesEndocrine System: Clarence L. Nuval, RMT, MDCla NuvalNo ratings yet
- Endocrine System - Life Sciences Questions and Answers - SanfoundryDocument8 pagesEndocrine System - Life Sciences Questions and Answers - SanfoundryHUAWEI HUAWEINo ratings yet
- Growth Hormone Responses To Sub-Maximal and Sprint Exercise: Keith StokesDocument14 pagesGrowth Hormone Responses To Sub-Maximal and Sprint Exercise: Keith StokesganduaffNo ratings yet
- Lesson 2 Endocrine SystemDocument18 pagesLesson 2 Endocrine SystemMARY JANE ANGELICA SEVANo ratings yet
- Anatomy and Physiology,: Lecture OutlineDocument79 pagesAnatomy and Physiology,: Lecture OutlineJorelyn Giron NaniongNo ratings yet