You might also like
- NanoencapsulationDocument61 pagesNanoencapsulationAbilash muraliNo ratings yet
- CSCL UltracentifugacionDocument4 pagesCSCL UltracentifugacionfebrauryNo ratings yet
- EM202257CON1014CHMDocument8 pagesEM202257CON1014CHMhoney arguellesNo ratings yet
- Articles: Optimization of Chitin Extraction From Shrimp ShellsDocument7 pagesArticles: Optimization of Chitin Extraction From Shrimp ShellsJuan Pablo EspinosaNo ratings yet
- Experimental Techniques For BioDocument6 pagesExperimental Techniques For BioMariam AsgharNo ratings yet
- Determination of The Mark-HouwinkDocument5 pagesDetermination of The Mark-HouwinkMariane AlcântaraNo ratings yet
- Amedica - Research PresentationDocument16 pagesAmedica - Research Presentationapi-343458772No ratings yet
- Mark Marlatt - Photosynthesis Lab and Report Track D - 5261224Document8 pagesMark Marlatt - Photosynthesis Lab and Report Track D - 5261224api-544304176No ratings yet
- Chlortetracycline, Oxytetracycline, and Tetracycline in Edible Animal TissuesDocument4 pagesChlortetracycline, Oxytetracycline, and Tetracycline in Edible Animal TissuesAhmed GwealyNo ratings yet
- Chemistry ExperimentDocument18 pagesChemistry ExperimentMalak MahmoudNo ratings yet
- MiMB Sample ChapterDocument10 pagesMiMB Sample ChapterSinivas ParthaNo ratings yet
- Recrystallization in Organic Chemistry LabDocument5 pagesRecrystallization in Organic Chemistry Labrc865No ratings yet
- IRIGANTI The Effects of Temperature On Sodium HypochloriteDocument4 pagesIRIGANTI The Effects of Temperature On Sodium HypochloritelupusebastianNo ratings yet
- Beetroot Core Practical Writing FrameDocument5 pagesBeetroot Core Practical Writing FrameJett0% (1)
- Grade 11-12 IB DP: Laboratory Manual For HL BiologyDocument128 pagesGrade 11-12 IB DP: Laboratory Manual For HL BiologyPhuc DangNo ratings yet
- 1041 FullDocument6 pages1041 FullHelen MejiaNo ratings yet
- Dialysate Circuit and Its Importance.: Sudhir BagaraoDocument49 pagesDialysate Circuit and Its Importance.: Sudhir BagaraoKinky DudeNo ratings yet
- Separation of Liquids by Fractional Distillation and Analysis by Gas ChromatographyDocument4 pagesSeparation of Liquids by Fractional Distillation and Analysis by Gas ChromatographyRuby RichiezNo ratings yet
- Holy Name University: Colloids Activity No. 2Document2 pagesHoly Name University: Colloids Activity No. 2Angelie Estilo Sarabia Paciente100% (1)
- Binary Phase Diagram of WaterDocument10 pagesBinary Phase Diagram of WaterkmkmNo ratings yet
- Osha pv2193Document9 pagesOsha pv2193luisalejandronovavNo ratings yet
- AOAC 994.10 - GC - Cholesterol in Foods PDFDocument2 pagesAOAC 994.10 - GC - Cholesterol in Foods PDFdennisaberliani100% (1)
- Stability of Andrographolide in Powdered AndrograpDocument5 pagesStability of Andrographolide in Powdered AndrograpNgô HưngNo ratings yet
- Chem87l Expt2 Padel PedarseDocument5 pagesChem87l Expt2 Padel Pedarsevince padelNo ratings yet
- Hayden Cassinelli - Photosynthesis Lab and Report Track C - 5258580Document6 pagesHayden Cassinelli - Photosynthesis Lab and Report Track C - 5258580api-548259020No ratings yet
- CDNA ExtractionDocument4 pagesCDNA Extractionvikashisar009No ratings yet
- Inhibition of Aldehyde Dehydrogenase IN Brain and Liver by CyanamideDocument5 pagesInhibition of Aldehyde Dehydrogenase IN Brain and Liver by CyanamideCristina Martín JiménezNo ratings yet
- Chem87l Expt5 Padel PedarseDocument5 pagesChem87l Expt5 Padel Pedarsevince padelNo ratings yet
- Fiber Aoac PDFDocument1 pageFiber Aoac PDFArellanes JmzNo ratings yet
- Preparation of A T-Butyl Chloride From T-Butyl Alcohol LabDocument5 pagesPreparation of A T-Butyl Chloride From T-Butyl Alcohol Labapi-548204552100% (1)
- Aoac978 10Document1 pageAoac978 10Mohammed ElsenduonyNo ratings yet
- tmpD44A TMPDocument15 pagestmpD44A TMPFrontiersNo ratings yet
- Wright 2019Document5 pagesWright 2019Raghav PratapNo ratings yet
- Batch Crystallizer: Rajesh Scientific IndustriesDocument8 pagesBatch Crystallizer: Rajesh Scientific IndustriesrajeshNo ratings yet
- Biomedical Electron Microscopy: Illustrated Methods and InterpretationsFrom EverandBiomedical Electron Microscopy: Illustrated Methods and InterpretationsNo ratings yet
- Elsevier Sequoia S. A., Lausanne A Kaddmiai Kiad6, BudapestDocument8 pagesElsevier Sequoia S. A., Lausanne A Kaddmiai Kiad6, Budapestgonzalo7cornejo7ramiNo ratings yet
- Vapour Liquid EquillibriumDocument15 pagesVapour Liquid EquillibriumMikho CingoNo ratings yet
- Exp't 41: The Reaction of Maleic Anhydride and CycloheptatrieneDocument5 pagesExp't 41: The Reaction of Maleic Anhydride and CycloheptatrienelovehopeNo ratings yet
- Batch Reactor ExpDocument12 pagesBatch Reactor ExpJack AndreasNo ratings yet
- Lab Report ChemisrtyDocument23 pagesLab Report ChemisrtyWakil NuckchadyNo ratings yet
- Section 02 Basic Tools and OperationsDocument45 pagesSection 02 Basic Tools and Operationslancelite88No ratings yet
- Chloride - Mercuric Nitrate Method PDFDocument6 pagesChloride - Mercuric Nitrate Method PDFYajNo ratings yet
- Metodo Hach 8206 para Cloruros Por Nitrato de MercurioDocument6 pagesMetodo Hach 8206 para Cloruros Por Nitrato de MercurioAIDEENo ratings yet
- 45.4.10 AOAC Official Method 994.10 Cholesterol in Foods: C) D) E) F) G) H)Document11 pages45.4.10 AOAC Official Method 994.10 Cholesterol in Foods: C) D) E) F) G) H)Angélica VargasNo ratings yet
- Report No 3 Quantitative Analysis A HydrateDocument7 pagesReport No 3 Quantitative Analysis A HydraterodneyperuNo ratings yet
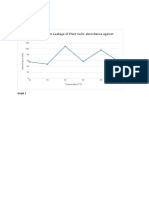
- Graph of Pigment Leakage of Plant Cells: Absorbance Against TemperaureDocument5 pagesGraph of Pigment Leakage of Plant Cells: Absorbance Against TemperaureZorina BiNo ratings yet
- REAGEN Doxycycline ELISA Test Kit ManualDocument11 pagesREAGEN Doxycycline ELISA Test Kit ManualpetertrungNo ratings yet
- Drying and Preservation of BacteriaDocument12 pagesDrying and Preservation of Bacteriateo20000No ratings yet
- Dimethoate Induced Changes in Serum Ca and Corpuscles of Stannius in Freshwater Catfish ExposureDocument5 pagesDimethoate Induced Changes in Serum Ca and Corpuscles of Stannius in Freshwater Catfish Exposurekni_zoologyNo ratings yet
- Profiling of Methyltransferases and Other S-Adenosyl - Homocysteine-Binding Proteins by Capture Compound Mass Spectrometry (CCMS)Document7 pagesProfiling of Methyltransferases and Other S-Adenosyl - Homocysteine-Binding Proteins by Capture Compound Mass Spectrometry (CCMS)Anh TranNo ratings yet
- The Influence of Lignin Degradation Products On Xylose Fermentation by Klebsiella PneumoniaeDocument4 pagesThe Influence of Lignin Degradation Products On Xylose Fermentation by Klebsiella Pneumoniaefilemon jaluNo ratings yet
- Chemy 310 Experiment 2Document5 pagesChemy 310 Experiment 2Faisal MumtazNo ratings yet
- Fermentation LabDocument12 pagesFermentation LabChiNo ratings yet
- AOAC Official Method 2003 - 01 Enumeration of Enterobaceriaceae in Selected FoodsDocument4 pagesAOAC Official Method 2003 - 01 Enumeration of Enterobaceriaceae in Selected FoodsaliciatapiaNo ratings yet
- Crystal Violet KineticsDocument9 pagesCrystal Violet KineticsMario VaNo ratings yet
- Lab Manual 2019Document27 pagesLab Manual 2019alexNo ratings yet
- To Increase in The Acticity and Selectivity in The Catalysis Via Surface Modification With Seld Assembled MonolayersDocument21 pagesTo Increase in The Acticity and Selectivity in The Catalysis Via Surface Modification With Seld Assembled Monolayerscute loverNo ratings yet
- Some Effects of Actinomycin D, Cycloheximide and Puromycin On Cell AdhesionDocument10 pagesSome Effects of Actinomycin D, Cycloheximide and Puromycin On Cell AdhesionFauziana NurhanisahNo ratings yet
- +the Journal of PathologyDocument7 pages+the Journal of PathologykeylapireNo ratings yet
- Plant Growth Regulator (Hormone)Document31 pagesPlant Growth Regulator (Hormone)Dilzar BasitNo ratings yet
- Ethylene Preparation and Its Application To PhysioDocument6 pagesEthylene Preparation and Its Application To PhysioDilzar BasitNo ratings yet
- Ethylene 180502141949Document29 pagesEthylene 180502141949Dilzar BasitNo ratings yet
- Springer Planta: This Content Downloaded From 130.193.211.143 On Mon, 24 Feb 2020 21:02:32 UTCDocument9 pagesSpringer Planta: This Content Downloaded From 130.193.211.143 On Mon, 24 Feb 2020 21:02:32 UTCDilzar BasitNo ratings yet
- Two Bioassay Techniques For Determining Abscisic Acid Concentrations1Document4 pagesTwo Bioassay Techniques For Determining Abscisic Acid Concentrations1Dilzar BasitNo ratings yet
- University of California, Los Angeles: PM AmDocument5 pagesUniversity of California, Los Angeles: PM AmDilzar BasitNo ratings yet
- ChromosomesDocument39 pagesChromosomesSuresh Kannan100% (1)
- As Biology AnswersDocument303 pagesAs Biology Answersprincejoe109No ratings yet
- Cell Division ActivityDocument4 pagesCell Division ActivityVivaMapwaNo ratings yet
- Notes - Chapter 3 - Cellular Level of OrganizationDocument7 pagesNotes - Chapter 3 - Cellular Level of OrganizationRogie P. BacosaNo ratings yet
- Genetics A Conceptual Approach 6th Edition Pierce Test BankDocument25 pagesGenetics A Conceptual Approach 6th Edition Pierce Test BankJackCoxmozj100% (56)
- Biology 30 Exemplars 2008Document85 pagesBiology 30 Exemplars 2008aaron93No ratings yet
- SR Elite & LTC Aits Grand Test - 17 Paper (25!04!2023)Document27 pagesSR Elite & LTC Aits Grand Test - 17 Paper (25!04!2023)vulurakashsharma2005No ratings yet
- 12th Biology Practicals 2022-23Document10 pages12th Biology Practicals 2022-23dig vijayNo ratings yet
- Chapter 19 - The Eukaryotic Cell CycleDocument47 pagesChapter 19 - The Eukaryotic Cell CycleRebecca Long HeiseNo ratings yet
- Cell Structure, Reproduction and DevelopmentDocument13 pagesCell Structure, Reproduction and Developmentmii chanrNo ratings yet
- Sci8 q4 Mod2 v4Document31 pagesSci8 q4 Mod2 v4Jay-ar RiosNo ratings yet
- Cells QuizDocument21 pagesCells QuizAj NavNo ratings yet
- Lecture Planner - Botany - Arjuna NEET 2025Document3 pagesLecture Planner - Botany - Arjuna NEET 2025Aaditya RathoreNo ratings yet
- Cell Cycle and Cell DivisionDocument8 pagesCell Cycle and Cell Divisionbhupeshgalani100% (2)
- Atlas of Antinuclear AntibodiesDocument40 pagesAtlas of Antinuclear AntibodiesFAIZAN KHANNo ratings yet
- S4 Mitotic Cell Division Lesson Plan PDFDocument4 pagesS4 Mitotic Cell Division Lesson Plan PDFShiella Mae Baltazar BulauitanNo ratings yet
- The Frequency GenerationDocument67 pagesThe Frequency Generationjayjonbeach100% (5)
- 2 Cell Cycle ChromosomesDocument12 pages2 Cell Cycle ChromosomesMallikarjuna MuthyaluNo ratings yet
- 211 Mitosis LabDocument5 pages211 Mitosis LabadekNo ratings yet
- Biophotonics & Light Communication - The Science Behind Energy HealingDocument25 pagesBiophotonics & Light Communication - The Science Behind Energy HealingIvan Roca100% (1)
- Lesson Plan Cell Division and MitosisDocument4 pagesLesson Plan Cell Division and MitosisEcha PakManNo ratings yet
- 8 Biotechnology ReviewerDocument3 pages8 Biotechnology ReviewerEunice Aruta100% (1)
- Biology 10th Edition Solomon Solutions Manual 1Document6 pagesBiology 10th Edition Solomon Solutions Manual 1donald100% (31)
- 4.1 - Cell Cycle Part 1Document5 pages4.1 - Cell Cycle Part 1Deomar Joseph ParadoNo ratings yet
- M13 Cell CycleDocument15 pagesM13 Cell CycleSamantha Kyle E. PedrigozaNo ratings yet
- Vocabulary PowerDocument5 pagesVocabulary PowerstrawhatalexNo ratings yet
- Chapter 20Document3 pagesChapter 20Shem Balaba Ramos SolivenNo ratings yet
- AnaPhy-lab ACTSDocument5 pagesAnaPhy-lab ACTSCEEJNo ratings yet
- Practical Manual Biol 0200 2021-2022Document39 pagesPractical Manual Biol 0200 2021-2022Anwaar SamaiNo ratings yet
- Biol 110 Essex Without Recitation Spring 2015Document7 pagesBiol 110 Essex Without Recitation Spring 2015api-281761748No ratings yet