You might also like
- INTP Parents - 16personalitiesDocument4 pagesINTP Parents - 16personalitiescelinelbNo ratings yet
- 1 s2.0 S0308814621025723 MainDocument12 pages1 s2.0 S0308814621025723 MainArthur RamadhaniNo ratings yet
- Evaluation of Physicochemical Properties of Trigona Sp. Stingless Bee Honey FromDocument9 pagesEvaluation of Physicochemical Properties of Trigona Sp. Stingless Bee Honey FromRobin WongNo ratings yet
- NizamDocument9 pagesNizamAmir MukmininNo ratings yet
- 8 fr-2018-106.r2 ZuwariahDocument7 pages8 fr-2018-106.r2 ZuwariahQuyên ĐàoNo ratings yet
- Journal of Food Composition and Analysis: Original Research ArticleDocument12 pagesJournal of Food Composition and Analysis: Original Research ArticleGabrielNo ratings yet
- KaracaliDocument9 pagesKaracaliProto TurkNo ratings yet
- LatviaResRuralDev 27th 2021-96-103Document9 pagesLatviaResRuralDev 27th 2021-96-103I-Simion GNo ratings yet
- 1 s2.0 S2405844023014093 MainDocument10 pages1 s2.0 S2405844023014093 Maingusmon sidikNo ratings yet
- Journal of Food Composition and Analysis: ReviewDocument12 pagesJournal of Food Composition and Analysis: ReviewWJ NgNo ratings yet
- Chemical Composition and Biological Activities of Beebread - ReviewDocument9 pagesChemical Composition and Biological Activities of Beebread - ReviewArJun AzisNo ratings yet
- LWT - Food Science and Technology: Damla Dag, Mete Kilercioglu, Mecit Halil OztopDocument9 pagesLWT - Food Science and Technology: Damla Dag, Mete Kilercioglu, Mecit Halil OztopAsri WidyasantiNo ratings yet
- FullPaperChickenFeetGelatinVol 30March2012BSJDocument14 pagesFullPaperChickenFeetGelatinVol 30March2012BSJSara KhaledNo ratings yet
- Trends in Food Science & TechnologyDocument13 pagesTrends in Food Science & TechnologyfrendystpNo ratings yet
- Gac (Momordica Cochinchinensis Spreng.) Fruit and Its Potentiality and Superiority In-Health BenefitsDocument9 pagesGac (Momordica Cochinchinensis Spreng.) Fruit and Its Potentiality and Superiority In-Health BenefitsEdgar BeltranNo ratings yet
- Archive of SID: Antimicrobial Peptides Derived From Goat's Milk Whey Proteins Obtained by Enzymatic HydrolysisDocument8 pagesArchive of SID: Antimicrobial Peptides Derived From Goat's Milk Whey Proteins Obtained by Enzymatic Hydrolysisredmi ferisNo ratings yet
- Article Peu Impo Isolation, Screening and Comprehensive Characterization of CandidateDocument9 pagesArticle Peu Impo Isolation, Screening and Comprehensive Characterization of CandidateMalak HamrouniNo ratings yet
- Characterization of Lipid ExtractsDocument8 pagesCharacterization of Lipid ExtractsCamila RochaNo ratings yet
- Comparison of Functional Properties of Edible Insects and Protein Preparations Thereof Ewelina Zieli Ska & Monika Kara & Barbara BaraniakDocument30 pagesComparison of Functional Properties of Edible Insects and Protein Preparations Thereof Ewelina Zieli Ska & Monika Kara & Barbara Baraniakbrent.lenis608100% (10)
- Protein Hidrolisat Antioksidan MaggotDocument8 pagesProtein Hidrolisat Antioksidan MaggotAmrul Ihsan ShinichiNo ratings yet
- Penerbit, 027Document9 pagesPenerbit, 027Hoong Wei JunNo ratings yet
- 1 s2.0 S0956713518300732 MainDocument59 pages1 s2.0 S0956713518300732 MainFelix WidjajaNo ratings yet
- The Antioxidant Effect of Mongolian Honey ProductsDocument6 pagesThe Antioxidant Effect of Mongolian Honey ProductsInternational Journal of Innovative Science and Research TechnologyNo ratings yet
- GlucanDocument5 pagesGlucanevan febriansyahNo ratings yet
- 1 s2.0 S0023643821000931 MainDocument8 pages1 s2.0 S0023643821000931 MainSally Salina FarahNo ratings yet
- Hassan Et Al., 2024Document15 pagesHassan Et Al., 2024Herda CahyaningrumNo ratings yet
- 1 s2.0 S2949824423001143 MainDocument12 pages1 s2.0 S2949824423001143 MainProyecto Orquídeas BPAMNo ratings yet
- Protein Isolates From Bambara Groundnut Voandz.Document19 pagesProtein Isolates From Bambara Groundnut Voandz.ilyas.tassineNo ratings yet
- Characterization and Comparative Study On StructuralDocument12 pagesCharacterization and Comparative Study On StructuralkaynnanathosNo ratings yet
- Screening Porcine DNA in Collagen Cream Cosmetic ProductsDocument6 pagesScreening Porcine DNA in Collagen Cream Cosmetic ProductsFarida Putri SyayarohNo ratings yet
- Antimicrobial Activity and Synergistic Effects of Honey and Propolis Produced by Apis Mellifera (European Honeybees)Document32 pagesAntimicrobial Activity and Synergistic Effects of Honey and Propolis Produced by Apis Mellifera (European Honeybees)Euler MendozaNo ratings yet
- Physicochemical, Antioxidant and Sensory Properties of Chocolate Spread Fortified With Jackfruit (Artocarpus Heterophyllus) FlourDocument9 pagesPhysicochemical, Antioxidant and Sensory Properties of Chocolate Spread Fortified With Jackfruit (Artocarpus Heterophyllus) FlourDisktirk12No ratings yet
- Honey Bee Research PaperDocument5 pagesHoney Bee Research Paperafnkwiytzcpszv100% (1)
- Optimization and Cholesterol Lowering Activity of Exopolysaccharide From Lactiplantibacillus Paraplantarum NCCP 962Document16 pagesOptimization and Cholesterol Lowering Activity of Exopolysaccharide From Lactiplantibacillus Paraplantarum NCCP 962nur rohmanNo ratings yet
- Potential Biological Activity of Acacia Honey: ArticleDocument8 pagesPotential Biological Activity of Acacia Honey: ArticleSayed Newaj ChowdhuryNo ratings yet
- The Antimicrobial Activity of Honey Against Common Equine Wound Bacterial IsolatesDocument5 pagesThe Antimicrobial Activity of Honey Against Common Equine Wound Bacterial IsolatesWilliam ChandlerNo ratings yet
- Evaluation of Nutritional Composition of Ripe Date Fruit (Phoenix Dactylifera L.) Pulp and Seed Grown in NigeriaDocument6 pagesEvaluation of Nutritional Composition of Ripe Date Fruit (Phoenix Dactylifera L.) Pulp and Seed Grown in NigeriaMamta AgarwalNo ratings yet
- Qualitative Evaluation of Honey Available in Bangladeshi MarketsDocument9 pagesQualitative Evaluation of Honey Available in Bangladeshi MarketsMehreen shehzadiNo ratings yet
- Molecules 26 01688 v3Document17 pagesMolecules 26 01688 v3keithNo ratings yet
- AntibioticsDocument8 pagesAntibioticsMedical UpdateNo ratings yet
- Antimicrobial Activity and Synergistic Effects of Honey and Propolis Produced by Apis Mellifera (European Honeybees) - 2Document36 pagesAntimicrobial Activity and Synergistic Effects of Honey and Propolis Produced by Apis Mellifera (European Honeybees) - 2Euler Mendoza100% (1)
- Honey CharacterizationDocument10 pagesHoney CharacterizationTen Percent ManNo ratings yet
- Arkan 2022Document8 pagesArkan 2022Briant TSNo ratings yet
- In Vitro Anti-Diabetic Activity of Stingless Bee Honey From Different BotanicalDocument6 pagesIn Vitro Anti-Diabetic Activity of Stingless Bee Honey From Different BotanicalpiangphetNo ratings yet
- Effect of Storage On Quality of Spirulina Snack BarsDocument8 pagesEffect of Storage On Quality of Spirulina Snack BarsIJAERS JOURNALNo ratings yet
- Food Bioscience: Didem S Ozeri Atik, Bas Ak Gürbüz, Esra B Olük, Ibrahim PalabıyıkDocument8 pagesFood Bioscience: Didem S Ozeri Atik, Bas Ak Gürbüz, Esra B Olük, Ibrahim Palabıyıkhasene keskinNo ratings yet
- Total Phenolic Contents and Colour Intensity of Malaysian Honeys From The Apis Spp. and Trigona Spp. BeesDocument6 pagesTotal Phenolic Contents and Colour Intensity of Malaysian Honeys From The Apis Spp. and Trigona Spp. BeesFajrinPramanaNo ratings yet
- 1 s2.0 S2666833521000277 MainDocument9 pages1 s2.0 S2666833521000277 Mainkhoanguyen2597No ratings yet
- Characterization of Banana Peel Pectin As Potential Halal Pharmaceutical ExcipientDocument12 pagesCharacterization of Banana Peel Pectin As Potential Halal Pharmaceutical ExcipientRobiatul AwwaliyahNo ratings yet
- Identification of Genes Involved in Shea Butter BiDocument11 pagesIdentification of Genes Involved in Shea Butter Binana sefaNo ratings yet
- 10 1016@j Foodcont 2020 107236 PDFDocument8 pages10 1016@j Foodcont 2020 107236 PDFvjvargas88No ratings yet
- StarFruit IJAR PDFDocument5 pagesStarFruit IJAR PDFtayyaba mehmoodNo ratings yet
- RJM FixDocument8 pagesRJM FixAbal khairNo ratings yet
- 1 s2.0 S0308814619305709 Main PDFDocument10 pages1 s2.0 S0308814619305709 Main PDFDorsophilaNo ratings yet
- Effects of Chromolaena Odorata Leaf Meal SupplemenDocument10 pagesEffects of Chromolaena Odorata Leaf Meal SupplemenEmmanuel OwonaNo ratings yet
- Redgwell2003 PDFDocument10 pagesRedgwell2003 PDFAna Sofía HozmanNo ratings yet
- Microbiological Quality of Set Yoghurt Supplemented With Turmeric Powder (Curcuma Longa) During StorageDocument7 pagesMicrobiological Quality of Set Yoghurt Supplemented With Turmeric Powder (Curcuma Longa) During StorageUmesh PoudelNo ratings yet
- A Comprehensive Review of Moringa Oleifera BioactiDocument18 pagesA Comprehensive Review of Moringa Oleifera BioactiMonserrat GonzalezNo ratings yet
- Gelatin Pig Skin MarketDocument7 pagesGelatin Pig Skin MarketRofiqSunaryantoNo ratings yet
- Sensorial MalvaviscoDocument6 pagesSensorial Malvavisco9cczpxjcmtNo ratings yet
- Extracellular Glycolipids of Yeasts: Biodiversity, Biochemistry, and ProspectsFrom EverandExtracellular Glycolipids of Yeasts: Biodiversity, Biochemistry, and ProspectsNo ratings yet
- PLC BasedDocument28 pagesPLC BasedJayven VillamaterNo ratings yet
- TFB - 2012 - Pollen Pellet Color Purity Identity A4 BookletDocument7 pagesTFB - 2012 - Pollen Pellet Color Purity Identity A4 BookletJayven VillamaterNo ratings yet
- Thesis Shanks JDocument307 pagesThesis Shanks JJayven VillamaterNo ratings yet
- ESAS Objectives 1Document15 pagesESAS Objectives 1Jayven VillamaterNo ratings yet
- Stingless Bees in Sunflower Pollination: Hemanth Kumar R, Srinivasa Reddy KM, Shishira D and Eshwarappa GDocument4 pagesStingless Bees in Sunflower Pollination: Hemanth Kumar R, Srinivasa Reddy KM, Shishira D and Eshwarappa GJayven VillamaterNo ratings yet
- Eebc 33223 3321l-Syllabus-2019Document11 pagesEebc 33223 3321l-Syllabus-2019Jayven VillamaterNo ratings yet
- PortfolioDocument26 pagesPortfolioJayven VillamaterNo ratings yet
- Datasheet MEC MPS200 v1 2018Document4 pagesDatasheet MEC MPS200 v1 2018Cepi Sindang KamulanNo ratings yet
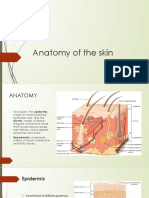
- Anatomy of The SkinDocument28 pagesAnatomy of The Skinay254No ratings yet
- Missoula County Fairgrounds Phase 2Document10 pagesMissoula County Fairgrounds Phase 2Olivia IversonNo ratings yet
- Scoop of Practice aOTADocument9 pagesScoop of Practice aOTAfercespedNo ratings yet
- Bitumen BasicsDocument25 pagesBitumen BasicsMILON KUMAR HORENo ratings yet
- Chapter Six Account Group General Fixed Assets Account Group (Gfaag)Document5 pagesChapter Six Account Group General Fixed Assets Account Group (Gfaag)meseleNo ratings yet
- Tractor Price and Speci Cations: Tractors in IndiaDocument4 pagesTractor Price and Speci Cations: Tractors in Indiatrupti kadamNo ratings yet
- L04-課文單片填空 (題目) (Day of the Dead)Document3 pagesL04-課文單片填空 (題目) (Day of the Dead)1020239korrnellNo ratings yet
- B - Cracked Tooth SyndromeDocument8 pagesB - Cracked Tooth SyndromeDavid TaylorNo ratings yet
- Prof. Madhavan - Ancient Wisdom of HealthDocument25 pagesProf. Madhavan - Ancient Wisdom of HealthProf. Madhavan100% (2)
- Hmo Details November 2022 1Document6 pagesHmo Details November 2022 1Saad BelloNo ratings yet
- Lecture 8 - ThermodynamicsDocument65 pagesLecture 8 - ThermodynamicsHasmaye PintoNo ratings yet
- Hospital - Data Collection & Literature StudyDocument42 pagesHospital - Data Collection & Literature StudyNagateja MallelaNo ratings yet
- Probni Test 1. Godina - Ina KlipaDocument4 pagesProbni Test 1. Godina - Ina KlipaMickoNo ratings yet
- Power Divider and Combiner: EE403-Microwave Engineering MTC, EE Dep., Electromagnetic Waves GroupDocument52 pagesPower Divider and Combiner: EE403-Microwave Engineering MTC, EE Dep., Electromagnetic Waves GroupHabibat El Rahman AshrafNo ratings yet
- NTJN, Full Conference Program - FINALDocument60 pagesNTJN, Full Conference Program - FINALtjprogramsNo ratings yet
- Building and Environment: Nabeel Ahmed Khan, Bishwajit BhattacharjeeDocument19 pagesBuilding and Environment: Nabeel Ahmed Khan, Bishwajit Bhattacharjeemercyella prasetyaNo ratings yet
- Resume Massage Therapist NtewDocument2 pagesResume Massage Therapist NtewPartheebanNo ratings yet
- Notice of Privacy Practices (Generic)Document3 pagesNotice of Privacy Practices (Generic)cecilled_08No ratings yet
- Chi - Square Test: PG Students: DR Amit Gujarathi DR Naresh GillDocument32 pagesChi - Square Test: PG Students: DR Amit Gujarathi DR Naresh GillNaresh GillNo ratings yet
- Ancon Tension Systems March 2008Document16 pagesAncon Tension Systems March 2008Slinky BillNo ratings yet
- Powerful Communication Tools For Successful Acupuncture PracticeDocument4 pagesPowerful Communication Tools For Successful Acupuncture Practicebinglei chenNo ratings yet
- Blood DonationDocument19 pagesBlood DonationsuruthiNo ratings yet
- Sol. Mock Test CBSE BiologyDocument3 pagesSol. Mock Test CBSE BiologysbarathiNo ratings yet
- Free Higher Education Application Form 1st Semester, SY 2021-2022Document1 pageFree Higher Education Application Form 1st Semester, SY 2021-2022Wheng NaragNo ratings yet
- Recommended Standards For Newborn ICU DesignDocument39 pagesRecommended Standards For Newborn ICU DesignAlbert SekarNo ratings yet
- G.R. No. 178741Document1 pageG.R. No. 178741Jefferson BagadiongNo ratings yet
- Astm d2729Document2 pagesAstm d2729Shan AdriasNo ratings yet