You might also like
- BMSC 230 Noted Chapter 2Document3 pagesBMSC 230 Noted Chapter 2shakshi dhruvNo ratings yet
- Theme 2: Included in This SectionDocument26 pagesTheme 2: Included in This SectionKate TaylorNo ratings yet
- Collection of Importance in MBCHMDocument7 pagesCollection of Importance in MBCHMAnonymous vCXYXdv45No ratings yet
- Glycolysis Regulation, Importance, and Fate of End ProductDocument7 pagesGlycolysis Regulation, Importance, and Fate of End ProductSulaiman AlmajeediNo ratings yet
- Selected Solutions To End of Chapter 15 ProblemsDocument4 pagesSelected Solutions To End of Chapter 15 ProblemsJeff D'erique Ozil TettehNo ratings yet
- Regulation of GluconeogenesisDocument2 pagesRegulation of GluconeogenesisAbbireddy SairamNo ratings yet
- BIO 202 Biochemistry II by Seyhun YURDUGÜL: Carbohydrate Metabolism II Regulation of GlycolysisDocument75 pagesBIO 202 Biochemistry II by Seyhun YURDUGÜL: Carbohydrate Metabolism II Regulation of GlycolysisYasin Çağrı KılıçerNo ratings yet
- Gluconeogenesis and Glycolysis Are Reciprocally RegulatedDocument3 pagesGluconeogenesis and Glycolysis Are Reciprocally Regulatedshakshi dhruvNo ratings yet
- Biochemistry: Glycogen MetabolismDocument78 pagesBiochemistry: Glycogen MetabolismJoseph YayenNo ratings yet
- Regulation of Glycogen MetabolismDocument18 pagesRegulation of Glycogen MetabolismNoel ManyiseNo ratings yet
- MBG312 Chp21Document48 pagesMBG312 Chp21Baran KirdarNo ratings yet
- Metabolisme GlikogenDocument17 pagesMetabolisme GlikogenMUHAMMAD RIDONo ratings yet
- Glycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryDocument25 pagesGlycolysis: Shekhar Chandra Yadav Lecturer Dept. of BiochemistryTULSI SHARMANo ratings yet
- Glycogenolysis: Gandham. RajeevDocument31 pagesGlycogenolysis: Gandham. RajeevVishesh DwivediNo ratings yet
- Metabolism & Glycolysis PAL - Raghda AdwanDocument74 pagesMetabolism & Glycolysis PAL - Raghda Adwanqueenmasa191No ratings yet
- GluconeogenesisDocument12 pagesGluconeogenesisRith Hengmeng (Study Smart)No ratings yet
- Biochemistry: Glycogen MetabolismDocument79 pagesBiochemistry: Glycogen Metabolismisabellacastro18No ratings yet
- Curs GluconeogenesisDocument25 pagesCurs GluconeogenesisOnofrei MariaNo ratings yet
- Control of Gluconeogenesis, Lecture 2Document47 pagesControl of Gluconeogenesis, Lecture 2China FonNo ratings yet
- GlycogenolysisDocument29 pagesGlycogenolysisAli AlqumaNo ratings yet
- Glycogen MetabolismDocument17 pagesGlycogen Metabolismmags wagsNo ratings yet
- Carbohydrate Storage& BreakdownDocument4 pagesCarbohydrate Storage& BreakdownMona DomingoNo ratings yet
- Lecture 11 CSU1123 Regulation of Glycolysis and GluconeogenesisDocument21 pagesLecture 11 CSU1123 Regulation of Glycolysis and Gluconeogenesischirag pandeyNo ratings yet
- Biochem Lec30Document4 pagesBiochem Lec30Louis FortunatoNo ratings yet
- GlycoDocument2 pagesGlycodeepankarborahNo ratings yet
- Gluco Neo GenesisDocument31 pagesGluco Neo GenesisJavedNo ratings yet
- Echevarria Jonille S. BSP 2Document6 pagesEchevarria Jonille S. BSP 2Jonille EchevarriaNo ratings yet
- GlycolysisDocument31 pagesGlycolysisrafea_naffa8326No ratings yet
- Chapter 14Document23 pagesChapter 14nhocnho_1993No ratings yet
- Dr. Tahir Mehmood: Bioenergetics and Metabolism (Cr.04) CHEM - 423 and 623Document43 pagesDr. Tahir Mehmood: Bioenergetics and Metabolism (Cr.04) CHEM - 423 and 623ASAD MALIKNo ratings yet
- Glycolysis:: The Central Pathway of Glucose DegradationDocument23 pagesGlycolysis:: The Central Pathway of Glucose DegradationMohammad Noman AkramNo ratings yet
- 2021 CHO Metabolism 1Document48 pages2021 CHO Metabolism 1xb2k9gzkc9No ratings yet
- Gluconeogenesis: The Pathway and RegulationDocument31 pagesGluconeogenesis: The Pathway and RegulationMunachande KanondoNo ratings yet
- Final Homework (20 Points) : Chloe Givens 4/17/16 Section: M 9:05amDocument3 pagesFinal Homework (20 Points) : Chloe Givens 4/17/16 Section: M 9:05amManupal YadavNo ratings yet
- Carbohydrate MetabolismDocument15 pagesCarbohydrate MetabolismPRIYANSHU KAUSHALNo ratings yet
- Carbohydrate Metabolism Glycolysis: Biochemistry 2 Code (4131)Document24 pagesCarbohydrate Metabolism Glycolysis: Biochemistry 2 Code (4131)essaNo ratings yet
- 4.metabolism of Glycogen: Biomedical ImportanceDocument9 pages4.metabolism of Glycogen: Biomedical ImportanceMd. Ismail HosenNo ratings yet
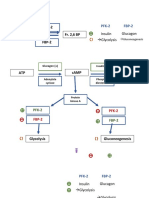
- F6P PFK-2 Fr. 2,6 BP FBP-2: Insulin Glucagon GlycolysisDocument4 pagesF6P PFK-2 Fr. 2,6 BP FBP-2: Insulin Glucagon GlycolysisKate TaylorNo ratings yet
- What Is MetabolismDocument10 pagesWhat Is MetabolismHoney Jane InutanNo ratings yet
- Metabolic Homeostasis: Regulation of Appetite, Energy Expenditure, Body Weight Unit-IVDocument26 pagesMetabolic Homeostasis: Regulation of Appetite, Energy Expenditure, Body Weight Unit-IVSherin SunnyNo ratings yet
- BiochemistryDocument30 pagesBiochemistryKriztine Mae GonzalesNo ratings yet
- Regulation of LipidsDocument13 pagesRegulation of LipidsNeha SrivastavaNo ratings yet
- Integración Metabólica y Vías UnidireccionalesDocument31 pagesIntegración Metabólica y Vías UnidireccionalesFabiola AmigoNo ratings yet
- Glucose Levels Maintaining MechanicsDocument7 pagesGlucose Levels Maintaining MechanicsSukhdeep KumarNo ratings yet
- GlycogenmetabDocument45 pagesGlycogenmetabNat TanNo ratings yet
- 2-Glycolysis RegulationDocument15 pages2-Glycolysis RegulationAbdul RaufNo ratings yet
- Glycolysis MergedDocument15 pagesGlycolysis MergedNikhitha NunnaNo ratings yet
- Biochemistry of Cardiovascular System: Agnes O. KwenangDocument70 pagesBiochemistry of Cardiovascular System: Agnes O. KwenangAdi Surya SyamNo ratings yet
- Principle of Metabolic RegulationDocument27 pagesPrinciple of Metabolic RegulationFlorentNo ratings yet
- GLYCOGENOLYISDocument14 pagesGLYCOGENOLYISshivam1092003meenaNo ratings yet
- Biochem CH 27 Integration of MetabolismDocument6 pagesBiochem CH 27 Integration of MetabolismSchat ZiNo ratings yet
- Biochem Lec ReviewerDocument25 pagesBiochem Lec ReviewerBarlaan Raniela Marie ReyesNo ratings yet
- 04 Carbohydrate 7-9 2022Document71 pages04 Carbohydrate 7-9 2022Azhar JirmandNo ratings yet
- Chapter 18 HandoutDocument8 pagesChapter 18 HandoutreynandNo ratings yet
- Carbohydrate MetabolismDocument57 pagesCarbohydrate MetabolismYulliza Kurniawaty L100% (1)
- Voet Energy MetabolismDocument19 pagesVoet Energy Metabolismanum0siddiqueNo ratings yet
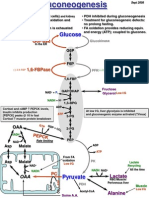
- #7 Gluconeogenesis - LA 2008Document2 pages#7 Gluconeogenesis - LA 2008bigoldtruck38No ratings yet
- Regulation of Glycogen MetabolismDocument36 pagesRegulation of Glycogen MetabolismNaji Mohamed AlfatihNo ratings yet
- Biochem Lec34Document3 pagesBiochem Lec34Louis FortunatoNo ratings yet
- Figure ADocument28 pagesFigure Atranquoctuan0204No ratings yet
- Figure 9.3Document1 pageFigure 9.3tranquoctuan0204No ratings yet
- CV Thanh Lap Hoi Dong Gui KhoaDocument1 pageCV Thanh Lap Hoi Dong Gui Khoatranquoctuan0204No ratings yet
- PDF Great Writing 3 Answer Key - CompressDocument16 pagesPDF Great Writing 3 Answer Key - Compresstranquoctuan0204No ratings yet
- GetDocument5 pagesGettranquoctuan0204No ratings yet
- GlobalDocument1 pageGlobaltranquoctuan0204No ratings yet
- NDIS Operational Procedures FBI 2005Document32 pagesNDIS Operational Procedures FBI 2005jac_blakeNo ratings yet
- FMDVDocument29 pagesFMDVmateus_laguardiaNo ratings yet
- Gel Electrophoresis Basics Worksheet: NameDocument2 pagesGel Electrophoresis Basics Worksheet: Nameapi-52265051475% (4)
- CRISPRDocument6 pagesCRISPRvijayjeoNo ratings yet
- Protein Synthesis Regulation ModelDocument3 pagesProtein Synthesis Regulation ModelzafiharoNo ratings yet
- Restriction Digest and Analysis of Lambda DNA Kit ManualDocument60 pagesRestriction Digest and Analysis of Lambda DNA Kit ManualJenny ReyesNo ratings yet
- Virology Journal: Predicting The Subcellular Localization of Viral Proteins Within A Mammalian Host CellDocument8 pagesVirology Journal: Predicting The Subcellular Localization of Viral Proteins Within A Mammalian Host CellRuy Lopez ClosedNo ratings yet
- Cyclins and Cyclin-Dependent Kinases (CDKS)Document16 pagesCyclins and Cyclin-Dependent Kinases (CDKS)Sudarshan KiniNo ratings yet
- February 2021 Teaching Roster - MBBSDocument4 pagesFebruary 2021 Teaching Roster - MBBSasjxhiBBniu01j2No ratings yet
- Proteomic Analysis ReviewDocument4 pagesProteomic Analysis ReviewWelzan SimonNo ratings yet
- Dna WsDocument1 pageDna WsLovryan Tadena AmilingNo ratings yet
- Kami Export - Erick Flesch - DNA-Replication-Overview-and-ReviewDocument2 pagesKami Export - Erick Flesch - DNA-Replication-Overview-and-ReviewFuckyouNo ratings yet
- Nucleic Acids WorksheetDocument4 pagesNucleic Acids WorksheetNatalie PembertonNo ratings yet
- Holt BiologyDocument30 pagesHolt BiologyKiro RemonNo ratings yet
- Unit 1Document15 pagesUnit 1Sakthivel PandianNo ratings yet
- Topic 1.3 Membrane StructureDocument4 pagesTopic 1.3 Membrane Structurejasmine wibawaNo ratings yet
- PP k2 Percubaan SBP 2007Document3 pagesPP k2 Percubaan SBP 2007kamil muhammadNo ratings yet
- Protein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm ViabilityDocument14 pagesProtein Signatures of Seminal Plasma From Bulls With Contrasting Frozen-Thawed Sperm Viabilitysaifulmangopo123No ratings yet
- Lesson 3 Biomolecules ReviewerDocument15 pagesLesson 3 Biomolecules Reviewerjohnromar VilasNo ratings yet
- Enzymes PDFDocument13 pagesEnzymes PDFPaul MarloweNo ratings yet
- Overview of Hemostasis - UpToDateDocument50 pagesOverview of Hemostasis - UpToDateTrung LêNo ratings yet
- Biology Dps - 112 Molecular Basis of Inheritance Biomentors Classes Online, Mumbai Date - 5 October 2018Document5 pagesBiology Dps - 112 Molecular Basis of Inheritance Biomentors Classes Online, Mumbai Date - 5 October 2018haida hamzaNo ratings yet
- Biochemistry 8th EditionDocument12 pagesBiochemistry 8th EditionxewazixNo ratings yet
- Aaaa PDFDocument8 pagesAaaa PDFgitanjali kashniNo ratings yet
- Genetics Problem SetDocument3 pagesGenetics Problem SetAakriti ThakurNo ratings yet
- Activity 1.3.1: Student Response Sheet: PART A-Restriction EnzymesDocument3 pagesActivity 1.3.1: Student Response Sheet: PART A-Restriction EnzymesRiley McCaffertyNo ratings yet
- Heat Coagulation TestDocument8 pagesHeat Coagulation TestRawan MaherNo ratings yet
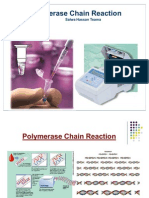
- Polymerase Chain Reaction: Salwa Hassan TeamaDocument56 pagesPolymerase Chain Reaction: Salwa Hassan TeamaSamiksha SharmaNo ratings yet
- Non-Nucleoside Inhibitors On NS5BDocument14 pagesNon-Nucleoside Inhibitors On NS5BmiklwlNo ratings yet
- Lehninger Ch26Document81 pagesLehninger Ch26AMAN KUMAR SINGH100% (1)