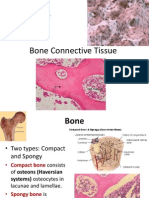
You might also like
- Bone Tissue-Growth and DevelopmentDocument53 pagesBone Tissue-Growth and Developmentgalaxyduck100% (1)
- BONEDocument48 pagesBONEnivethaseshaNo ratings yet
- HANDOUTS Prelim CH 6Document40 pagesHANDOUTS Prelim CH 6Abia Annieson A. LorenzoNo ratings yet
- Chapter 6 Content Review Questions 1-8Document3 pagesChapter 6 Content Review Questions 1-8Rhonique MorganNo ratings yet
- Human Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFDocument28 pagesHuman Anatomy 5Th Edition Saladin Solutions Manual Full Chapter PDFJenniferWhitebctr100% (10)
- Human Anatomy 5th Edition Saladin Solutions ManualDocument7 pagesHuman Anatomy 5th Edition Saladin Solutions Manualjillhenrysetcjnzrfp100% (25)
- Chapter 6 MariebDocument6 pagesChapter 6 Mariebmissy23papNo ratings yet
- 2) Development and Growth of The BonesDocument3 pages2) Development and Growth of The BonesAris PaparisNo ratings yet
- FRACTURESDocument85 pagesFRACTURESSteven OdhiamboNo ratings yet
- Bone As A Living Dynamic TissueDocument14 pagesBone As A Living Dynamic TissueSuraj_Subedi100% (6)
- OssificationDocument3 pagesOssificationFatin Nabihah JamilNo ratings yet
- Cartilage & BoneDocument8 pagesCartilage & BoneRehab OmerNo ratings yet
- BoneDocument3 pagesBonePeter 'Pierre' RobsonNo ratings yet
- Anaphy AnalysisDocument6 pagesAnaphy AnalysisLuna ScorpiusNo ratings yet
- 05 Locomotor SkeletalDocument288 pages05 Locomotor SkeletalkalumbaaquilaNo ratings yet
- Chapter 6 Bone Tissue 2304Document37 pagesChapter 6 Bone Tissue 2304Sav Oli100% (1)
- Human Anatomy 4th Edition Saladin Solutions Manual 1Document36 pagesHuman Anatomy 4th Edition Saladin Solutions Manual 1donnawugnwsjrzcxt100% (21)
- Human Anatomy 4Th Edition Saladin Solutions Manual Full Chapter PDFDocument27 pagesHuman Anatomy 4Th Edition Saladin Solutions Manual Full Chapter PDFdora.ivy892100% (16)
- Skeletal System: CartilageDocument20 pagesSkeletal System: CartilageGrace PascualNo ratings yet
- Alveolar BoneDocument114 pagesAlveolar BoneDent DealsNo ratings yet
- Dev Skeletal SystemR2023Document83 pagesDev Skeletal SystemR2023amandagnaw19No ratings yet
- SKELETAL SYSTEM ReviewerDocument7 pagesSKELETAL SYSTEM ReviewerKrize Colene dela CruzNo ratings yet
- Practical Work No. 7: Faculty of MedicineDocument3 pagesPractical Work No. 7: Faculty of MedicineAndrei TimofteNo ratings yet
- LP 7 Connective Tissue BoneDocument3 pagesLP 7 Connective Tissue BoneAndrei TimofteNo ratings yet
- BC (H) - IV-Human Physiology-Bone Structure and Formation-2 & 3Document32 pagesBC (H) - IV-Human Physiology-Bone Structure and Formation-2 & 3Starco ForeverNo ratings yet
- BoneDocument33 pagesBoneNihal BilalNo ratings yet
- 1 Bone Tissue 2021-22Document35 pages1 Bone Tissue 2021-22Ali AfifNo ratings yet
- Lecture: Histology of Cartilage and BoneDocument49 pagesLecture: Histology of Cartilage and BonevictorNo ratings yet
- Skeletal System Lesson 2Document40 pagesSkeletal System Lesson 2Ella Nika FangonNo ratings yet
- Histogenesis of BoneDocument6 pagesHistogenesis of BoneAlya Putri KhairaniNo ratings yet
- Tolentino, Edjey Week4histolectDocument5 pagesTolentino, Edjey Week4histolectEDJEY TOLENTINONo ratings yet
- Functions of BoneDocument7 pagesFunctions of BoneDTM FHKNo ratings yet
- Ossification of Bone.Document22 pagesOssification of Bone.Shimmering MoonNo ratings yet
- CHAPTER 6 - Skeletal SystemDocument4 pagesCHAPTER 6 - Skeletal SystemArlen Joy V. AMPARONo ratings yet
- The Skeletal System HDocument23 pagesThe Skeletal System HRajorshi MishraNo ratings yet
- Bone Development and Growth: Rosy Setiawati and Paulus RahardjoDocument20 pagesBone Development and Growth: Rosy Setiawati and Paulus Rahardjoputri aisheNo ratings yet
- ALVEOLAR BoneDocument72 pagesALVEOLAR BoneArchana50% (2)
- m5 Skeletal SystemDocument33 pagesm5 Skeletal Systemapi-464344582No ratings yet
- Bone & Ossification: Histology Part: Module2, Unit 3Document14 pagesBone & Ossification: Histology Part: Module2, Unit 3Obansa John100% (1)
- DO - DPM Bone Formation D2L 2014Document7 pagesDO - DPM Bone Formation D2L 2014snlee06No ratings yet
- Lecture: Histology of Cartilage and BoneDocument49 pagesLecture: Histology of Cartilage and BoneStreet Fhigteer AgitatorNo ratings yet
- Bone and CartilageDocument56 pagesBone and CartilageGodwin ZengeniNo ratings yet
- BONE DEVELOPMENT XDocument5 pagesBONE DEVELOPMENT XabdulNo ratings yet
- Bone & Cartilage Review QuestionsDocument5 pagesBone & Cartilage Review QuestionsTaonga ZuluNo ratings yet
- Cartilage: Histology Part 1: Module2, Unit 2Document30 pagesCartilage: Histology Part 1: Module2, Unit 2Obansa JohnNo ratings yet
- Intramembranous Ossification Involves (Meliputi) The ReplacementDocument8 pagesIntramembranous Ossification Involves (Meliputi) The ReplacementMuhammad SaifullohNo ratings yet
- 1.2 Cartilage and BoneDocument27 pages1.2 Cartilage and BonePema GaleyNo ratings yet
- Skeletal SystemDocument200 pagesSkeletal SystemJB AriasdcNo ratings yet
- Bone Physiology: By: Sandesh. A. PDocument75 pagesBone Physiology: By: Sandesh. A. PAnalhaq ShaikhNo ratings yet
- BSN1 Unit 5 Skeletal SystemDocument101 pagesBSN1 Unit 5 Skeletal SystemCherub Chua100% (1)
- Anatomy and Physiology Musculoskeletal System Chap 6 - 2401Document18 pagesAnatomy and Physiology Musculoskeletal System Chap 6 - 2401slyfoxkittyNo ratings yet
- Anatomy and PhysiologyDocument26 pagesAnatomy and Physiologymusondaaugustine07_4No ratings yet
- 02 Skeletal SystemDocument32 pages02 Skeletal SystemHarshika KDGNo ratings yet
- Alv - Bone FinalDocument28 pagesAlv - Bone Finalv.shivakumarNo ratings yet
- Chapter 6 Cartilage and BoneDocument5 pagesChapter 6 Cartilage and BoneSuperjunior8No ratings yet
- Reviewer Zoo LecDocument27 pagesReviewer Zoo LecayeyedumpNo ratings yet
- Module-5-The Skeletal SystemDocument13 pagesModule-5-The Skeletal SystemMARIA CORAZON CONTANTENo ratings yet
- O Is Composed of Bones, Cartilage, and Ligaments o CartilageDocument13 pagesO Is Composed of Bones, Cartilage, and Ligaments o CartilageVarel JoanNo ratings yet
- Advanced farriery knowledge: A study guide and AWCF theory course companionFrom EverandAdvanced farriery knowledge: A study guide and AWCF theory course companionNo ratings yet
- (Supportive Tissue) - (DVM 1st Semester) - (Morning) - (A Section)Document10 pages(Supportive Tissue) - (DVM 1st Semester) - (Morning) - (A Section)hely shahNo ratings yet
- Repair of Full Thickness Articular Cartilage Defects Using IEIK13 Self Assembling Peptide Hydrogel in A Non Human Primate ModelDocument17 pagesRepair of Full Thickness Articular Cartilage Defects Using IEIK13 Self Assembling Peptide Hydrogel in A Non Human Primate Model9147. 5125'321199316No ratings yet
- BBA - Molecular Basis of Disease: ReviewDocument9 pagesBBA - Molecular Basis of Disease: ReviewAbdulaziz AlanziNo ratings yet
- Pathophysiology of OsteoarthritisDocument3 pagesPathophysiology of OsteoarthritisHajar ToharNo ratings yet
- Connective TissueDocument202 pagesConnective TissuePeter ThompsonNo ratings yet
- Connective Tissue NotesDocument6 pagesConnective Tissue Notesferdinand padillaNo ratings yet
- Strategies To Convert Cells Into Hyaline Cartilage: Magic Spells For Adult Stem CellsDocument28 pagesStrategies To Convert Cells Into Hyaline Cartilage: Magic Spells For Adult Stem CellsMahmood-S ChoudheryNo ratings yet
- Asterion Practical Handbook of Anatomy-2eDocument265 pagesAsterion Practical Handbook of Anatomy-2eMohammad AkhtarNo ratings yet
- Histology of The Cartilage (Week 7)Document8 pagesHistology of The Cartilage (Week 7)Robaina WisNo ratings yet
- Bone Tumors in Domestic Animals, Comparative Clinical Pathology (VetBooks - Ir)Document207 pagesBone Tumors in Domestic Animals, Comparative Clinical Pathology (VetBooks - Ir)fisheirNo ratings yet
- Condylar CartilageDocument11 pagesCondylar CartilageMahesh kumarNo ratings yet
- Chapter 54 - Pathogenesis of OsteochondrosisDocument9 pagesChapter 54 - Pathogenesis of OsteochondrosisRam RamNo ratings yet
- Articular Cartilage - Basic Science - OrthobulletsDocument6 pagesArticular Cartilage - Basic Science - OrthobulletsWina SusanaNo ratings yet
- Final!Document14 pagesFinal!JoshuaNo ratings yet
- Osteoarthritis PathopysiologyDocument4 pagesOsteoarthritis PathopysiologySolsona Natl HS MaanantengNo ratings yet
- CROBM 1996 Kieswetter 329 45Document17 pagesCROBM 1996 Kieswetter 329 45Jontavius GilbertNo ratings yet
- Biological Approaches For Cartilage RepairDocument9 pagesBiological Approaches For Cartilage RepairPoppyA.NamiraNo ratings yet
- Platelet Rich Plasma For Bone Healing and RegenerationDocument21 pagesPlatelet Rich Plasma For Bone Healing and RegenerationAnonymous Yr9JXHYpPANo ratings yet
- Anti Inflammatory and Analgesic Activity ofDocument172 pagesAnti Inflammatory and Analgesic Activity ofR.G.ManikarajanNo ratings yet
- Mats Brittberg - Konrad Slynarski - Lower Extremity Joint Preservation - Techniques For Treating The Hip, Knee, and Ankle-Springer (2021)Document340 pagesMats Brittberg - Konrad Slynarski - Lower Extremity Joint Preservation - Techniques For Treating The Hip, Knee, and Ankle-Springer (2021)Renan Sequini FavaroNo ratings yet
- Bài Báo 3Document10 pagesBài Báo 3HoangNgocAnhNhanNo ratings yet
- Human Histology 7 CartilageDocument6 pagesHuman Histology 7 CartilageBlubby BleuNo ratings yet
- Stem Cells and The Future of Dental CareDocument5 pagesStem Cells and The Future of Dental CarearisNo ratings yet
- Osteoarthritis: Epidemiology, Risk Factors, and PathophysiologyDocument10 pagesOsteoarthritis: Epidemiology, Risk Factors, and PathophysiologyMaryama AflahaNo ratings yet
- Histo CompilationDocument217 pagesHisto CompilationNico LokoNo ratings yet
- Developmental Biology Final Exam ReviewDocument12 pagesDevelopmental Biology Final Exam ReviewB. S.No ratings yet
- Histopathology Diagnosis PDFDocument231 pagesHistopathology Diagnosis PDFMarvi Umair100% (2)
- Bone and CartilageDocument56 pagesBone and CartilageGodwin ZengeniNo ratings yet
- Bone FormationDocument26 pagesBone FormationNmesomaNo ratings yet
- Histology-World! Histology Testbank-Cartilage 2aDocument7 pagesHistology-World! Histology Testbank-Cartilage 2aAya AhmedNo ratings yet