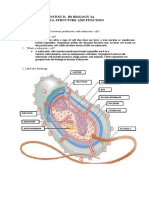
You might also like
- The Chlamydomonas Sourcebook: A Comprehensive Guide to Biology and Laboratory UseFrom EverandThe Chlamydomonas Sourcebook: A Comprehensive Guide to Biology and Laboratory UseNo ratings yet
- Cell The Unit of Life NotesDocument5 pagesCell The Unit of Life Notesk8392188No ratings yet
- Chloroplast EvolutionDocument3 pagesChloroplast EvolutionobaytutNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Class Notes PersonalDocument2 pagesClass Notes PersonalLau FernandezNo ratings yet
- Microbial Genetics: Extra-Chromosomal InheritanceDocument24 pagesMicrobial Genetics: Extra-Chromosomal Inheritancearlene eranNo ratings yet
- GENBIO - MEIOSIS II (Group 2)Document17 pagesGENBIO - MEIOSIS II (Group 2)Alyssa Sofia LofamiaNo ratings yet
- Practice Exam Question Practice Exam QuestionDocument5 pagesPractice Exam Question Practice Exam QuestionLily RobinsonNo ratings yet
- Mitosis Vs MeiosisDocument7 pagesMitosis Vs MeiosisJ.L.No ratings yet
- Developmental Biology 466Document4 pagesDevelopmental Biology 466Pearl PascuaNo ratings yet
- 745 MycoplasmaDocument79 pages745 MycoplasmadrbhaveshparmarNo ratings yet
- MCB 307Document2 pagesMCB 307Emmanuel ogenyiNo ratings yet
- 1.6 Cell Division Skeleton NotesDocument15 pages1.6 Cell Division Skeleton Notesadri baigorriNo ratings yet
- Cell Division: Dr. P. Pokorski University of Michigan - Dearborn Department of BiologyDocument52 pagesCell Division: Dr. P. Pokorski University of Michigan - Dearborn Department of BiologyZuha HarithNo ratings yet
- Chapter 12 - The Cell CycleDocument2 pagesChapter 12 - The Cell CycleAlexa MacailoNo ratings yet
- Special ChromosomesDocument68 pagesSpecial ChromosomesRashmi AshokNo ratings yet
- MODULE 4-GeneticsDocument14 pagesMODULE 4-GeneticsDivine Grace CincoNo ratings yet
- General Biology 1 Second Quarter: LearningDocument4 pagesGeneral Biology 1 Second Quarter: Learningcheryl galletoNo ratings yet
- Polytene ChromosomesDocument6 pagesPolytene ChromosomesKhush BakhtNo ratings yet
- Biology HL - ANSWERS - Second Edition - Pearson 2014 PDFDocument73 pagesBiology HL - ANSWERS - Second Edition - Pearson 2014 PDFAngela LinNo ratings yet
- Agata1999 PDFDocument7 pagesAgata1999 PDFMARIA CAMILA VELASCO CASTAÑEDANo ratings yet
- RBalderas - CBE3 Pre Test Answer KeyDocument39 pagesRBalderas - CBE3 Pre Test Answer KeyMikyle NatorNo ratings yet
- Haploidy PolyploidyDocument15 pagesHaploidy Polyploidyshagun brarNo ratings yet
- The Biochemistry of Malaria: Yongyuth Yuthavong and Prapon WilairatDocument6 pagesThe Biochemistry of Malaria: Yongyuth Yuthavong and Prapon Wilairataliyu belloNo ratings yet
- UntitledDocument88 pagesUntitledBamboo CaneNo ratings yet
- CelldivisionDocument52 pagesCelldivisionDrShafici Osman AdanNo ratings yet
- Biology: What Is Living Organisms?Document13 pagesBiology: What Is Living Organisms?Rohit SharmaNo ratings yet
- Exercise 5Document5 pagesExercise 5holie madroneroNo ratings yet
- Mitosis Worksheet & Diagram IdentificationDocument3 pagesMitosis Worksheet & Diagram IdentificationTyrese SmithNo ratings yet
- Gen Bio ReviewerDocument5 pagesGen Bio ReviewerJamon Ceballos Earl JensenNo ratings yet
- 5 - MeiosisDocument52 pages5 - MeiosissalmaNo ratings yet
- Biology 11th 14 - 01 - 2024 - 001525Document9 pagesBiology 11th 14 - 01 - 2024 - 001525zabiu8926No ratings yet
- Hybridomatechnology 190930140737Document20 pagesHybridomatechnology 190930140737Raluca IoanaNo ratings yet
- 331 FullDocument10 pages331 FullJean Pierre Chastre LuzaNo ratings yet
- MeiosisDocument30 pagesMeiosisngulubetsereletso89No ratings yet
- Chapter16 PDFDocument21 pagesChapter16 PDFsgw67No ratings yet
- Aneuploidy Cleavage Furrow and Cell Plate Karyotype: Distinguish BetweenDocument2 pagesAneuploidy Cleavage Furrow and Cell Plate Karyotype: Distinguish Betweeneclipse1130No ratings yet
- Grade 9 - Second Quarter SCIENCE by REWARD FULGUERASDocument4 pagesGrade 9 - Second Quarter SCIENCE by REWARD FULGUERASrewardfulgueras9No ratings yet
- IB Biology Notes - 42 MeiosisDocument7 pagesIB Biology Notes - 42 MeiosisJohn Philip D. NapalNo ratings yet
- AP Bio Chapter 13 NotesDocument5 pagesAP Bio Chapter 13 NotesAndrew SongNo ratings yet
- CBSE Quick Revision Notes (Class-11 Biology) Chapter-10 Cell Cycle and Cell DivisionDocument4 pagesCBSE Quick Revision Notes (Class-11 Biology) Chapter-10 Cell Cycle and Cell DivisionArnavNo ratings yet
- BIOL 3121 - Lecture 13 - 081116Document91 pagesBIOL 3121 - Lecture 13 - 081116Sara HuebnerNo ratings yet
- Cell ReproductionDocument84 pagesCell ReproductionCheska OlanamNo ratings yet
- About Pleomorphism. Suppressed Medical KnowledgeDocument17 pagesAbout Pleomorphism. Suppressed Medical KnowledgeCarl MacCord50% (2)
- Significance or Applications of Mitosis/Meiosis: General Biology 1Document26 pagesSignificance or Applications of Mitosis/Meiosis: General Biology 1beaNo ratings yet
- SIK2006 Lecture 02 Selected SlidesDocument30 pagesSIK2006 Lecture 02 Selected SlidesIIV190014 STUDENTNo ratings yet
- Pembelahan SelDocument53 pagesPembelahan SelArmi Ikawati ZainabNo ratings yet
- U Tube ExperimentDocument143 pagesU Tube ExperimentAbhishek SinghNo ratings yet
- Cell Cycle and Cell DivisionDocument6 pagesCell Cycle and Cell DivisionJeyanthiNo ratings yet
- GeneticsDocument29 pagesGeneticskmakabe78No ratings yet
- 8 Cell Structure and FunctionDocument21 pages8 Cell Structure and FunctionVince RetutaNo ratings yet
- Normoblastic Maturation: ErythropoiesisDocument2 pagesNormoblastic Maturation: ErythropoiesisKenneth Jake BatiduanNo ratings yet
- Gen BioDocument5 pagesGen BioDanilo JR. BuaquiñaNo ratings yet
- Plant Cell E-BookDocument21 pagesPlant Cell E-BookSammy ShnayderNo ratings yet
- Biol Activity 5 Word2003Document16 pagesBiol Activity 5 Word2003Faith Jessica ParanNo ratings yet
- ALTIUS - Unit Test # 2Document11 pagesALTIUS - Unit Test # 2Avrajoty RoyNo ratings yet
- In Liquid Medium Colchicine Treatment Induces Non Chimerical Doubled-Diploids in A Wide Range of Mono-And Interspecific Diploid Banana ClonesDocument10 pagesIn Liquid Medium Colchicine Treatment Induces Non Chimerical Doubled-Diploids in A Wide Range of Mono-And Interspecific Diploid Banana ClonesKarla ReyesNo ratings yet
- Genetics - Chapter 2 - Mitosis and Meiosis-Cellular ReproductionDocument37 pagesGenetics - Chapter 2 - Mitosis and Meiosis-Cellular ReproductionDuy AnhNo ratings yet
- 12th Bio Botany Study Material English Medium PDFDocument26 pages12th Bio Botany Study Material English Medium PDFBASKARANNo ratings yet
- Agargel PropertiesDocument3 pagesAgargel PropertiesLászló SágiNo ratings yet
- SterikonDocument4 pagesSterikonLászló SágiNo ratings yet
- Druart 1993Document3 pagesDruart 1993László SágiNo ratings yet
- 15 Minute Beginner Stretching Routine (Tom Merrick)Document4 pages15 Minute Beginner Stretching Routine (Tom Merrick)László SágiNo ratings yet
- Skirvin 1986Document3 pagesSkirvin 1986László SágiNo ratings yet
- Srivastava 2018Document25 pagesSrivastava 2018László SágiNo ratings yet
- Sawilowsky 2002Document13 pagesSawilowsky 2002László SágiNo ratings yet
- Sunderland 1977Document3 pagesSunderland 1977László SágiNo ratings yet
- Rawson 2011Document14 pagesRawson 2011László SágiNo ratings yet
- Wenzel 1975Document5 pagesWenzel 1975László SágiNo ratings yet
- Zargar 2022Document21 pagesZargar 2022László SágiNo ratings yet
- Chase 1964Document7 pagesChase 1964László SágiNo ratings yet
- Yang 2021Document11 pagesYang 2021László SágiNo ratings yet
- Bottanelli 2018Document7 pagesBottanelli 2018László SágiNo ratings yet
- Wei 1982Document3 pagesWei 1982László SágiNo ratings yet
- Cramer 2019Document11 pagesCramer 2019László SágiNo ratings yet
- Bian 2018Document16 pagesBian 2018László SágiNo ratings yet
- Tang 2011Document17 pagesTang 2011László SágiNo ratings yet
- Nejat 2017Document16 pagesNejat 2017László SágiNo ratings yet
- Groth 2012Document21 pagesGroth 2012László SágiNo ratings yet
- Khaled ThesisDocument119 pagesKhaled ThesisLászló SágiNo ratings yet
- Glenn 2013Document17 pagesGlenn 2013László SágiNo ratings yet
- Yao 2023Document16 pagesYao 2023László SágiNo ratings yet
- ESM 244 Lecture 4Document39 pagesESM 244 Lecture 4László SágiNo ratings yet
- ESM 244 Lecture 3Document34 pagesESM 244 Lecture 3László SágiNo ratings yet
- Chase 1963Document7 pagesChase 1963László SágiNo ratings yet
- Chase 1969 ADocument52 pagesChase 1969 ALászló SágiNo ratings yet
- Gniazdowski 2017Document24 pagesGniazdowski 2017László SágiNo ratings yet
- Chase 1949 ADocument5 pagesChase 1949 ALászló SágiNo ratings yet
- Visscher PresentDocument36 pagesVisscher PresentLászló SágiNo ratings yet
- Faults and EarthquakesDocument32 pagesFaults and EarthquakesArdoKimayKimArdoNo ratings yet
- STUDIOBRICKS QUOTE ONE ONE PLUS Automized Quote10Document38 pagesSTUDIOBRICKS QUOTE ONE ONE PLUS Automized Quote10Brady WilliamsNo ratings yet
- Algebra Adventures WorkbookDocument43 pagesAlgebra Adventures Workbookbuzuleacnadya100% (3)
- Bechelor ThesisDocument71 pagesBechelor ThesisLê KiênNo ratings yet
- Lesson Plan Monster GeneticsDocument2 pagesLesson Plan Monster Geneticsapi-312959802No ratings yet
- Fluido Barrera SeleccionDocument20 pagesFluido Barrera Seleccionmartin.rubenNo ratings yet
- Vital Signs 10 Am 2pm Clinical Interpretation: 37.0 Degrees Celsius 37.5 Degrees CelsiusDocument4 pagesVital Signs 10 Am 2pm Clinical Interpretation: 37.0 Degrees Celsius 37.5 Degrees CelsiusYamete KudasaiNo ratings yet
- IEDA 2010 Assignment 2 - SolutionDocument5 pagesIEDA 2010 Assignment 2 - SolutionNorris wongNo ratings yet
- Torrico ReinforcedConcrete 01 PDFDocument8 pagesTorrico ReinforcedConcrete 01 PDFNajib A. CasanNo ratings yet
- Diesel Power Plant Operation and Maintenance NC IIDocument64 pagesDiesel Power Plant Operation and Maintenance NC IIMonica D'gorgeous100% (2)
- PCT - 1351913 Eatnon ManualDocument326 pagesPCT - 1351913 Eatnon Manualmartha BernalNo ratings yet
- Canon Eos 40D Service Manual Repair GuideDocument209 pagesCanon Eos 40D Service Manual Repair Guidezstarce100% (1)
- METU ENVE202 Laboratory ManualDocument47 pagesMETU ENVE202 Laboratory ManualSarp ÇelebiNo ratings yet
- Non-Enzymatic Glycosylation and Pharmaceutical Intervention: Monica MorganDocument44 pagesNon-Enzymatic Glycosylation and Pharmaceutical Intervention: Monica MorganYeraldith RojasNo ratings yet
- Yoga For Problems of EyesDocument52 pagesYoga For Problems of EyesArun Thulasi100% (2)
- Solution Manual For Essentials of Chemical Reaction Engineering by FoglerDocument24 pagesSolution Manual For Essentials of Chemical Reaction Engineering by FoglerCecelia Sams100% (34)
- Benefits of Aviation Brazil 2011 IATADocument28 pagesBenefits of Aviation Brazil 2011 IATAKatia Mantovani MirandaNo ratings yet
- Tremulus Playbook Set IV - The PilotDocument2 pagesTremulus Playbook Set IV - The Pilotandrewh3No ratings yet
- Biology Today: Systematics of Marine Intervertebrates Evaluating Dr. Ken HalanychDocument4 pagesBiology Today: Systematics of Marine Intervertebrates Evaluating Dr. Ken Halanychapi-340402818No ratings yet
- Presentation On 3 Phase MotorDocument14 pagesPresentation On 3 Phase MotorPushkar PanditNo ratings yet
- Instruction Manual Professional: Digital Control Unit Cem7Document98 pagesInstruction Manual Professional: Digital Control Unit Cem7sonnguyen273100% (1)
- State Space Models: (Valid For Any Nonlinear Causal System)Document59 pagesState Space Models: (Valid For Any Nonlinear Causal System)Anonymous HsoXPyNo ratings yet
- Installation and Operation Manual: 14,000 Pound Capacity Commercial Grade Four-Post Lifts Models: HD-14TDocument52 pagesInstallation and Operation Manual: 14,000 Pound Capacity Commercial Grade Four-Post Lifts Models: HD-14TLê Xuân SangNo ratings yet
- Drill Stem TestDocument3 pagesDrill Stem Testpoliskarma0% (1)
- The Story of KeeshDocument4 pagesThe Story of KeeshEmman SanoNo ratings yet
- Guru Nadi - Full 3 PartsDocument35 pagesGuru Nadi - Full 3 Partspm plassanalNo ratings yet
- Lab 4 Enzymes and The Catalase LabDocument18 pagesLab 4 Enzymes and The Catalase LabAmy Hollingsworth100% (1)
- Honey: A Reservoir For Microorganisms and An Inhibitory Agent For MicrobesDocument7 pagesHoney: A Reservoir For Microorganisms and An Inhibitory Agent For MicrobesVidianka RembulanNo ratings yet
- Exercise 1 - TVM & Equivalence 2.0Document5 pagesExercise 1 - TVM & Equivalence 2.0Bayu PurnamaNo ratings yet
- Dell PowerEdge R210II TechnicalGuideDocument57 pagesDell PowerEdge R210II TechnicalGuideksypolitNo ratings yet