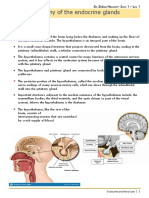
You might also like
- Lab Exercise 2Document4 pagesLab Exercise 2Angela Robles0% (1)
- HLTAAP001 Recognize Healthy Body System 9Document17 pagesHLTAAP001 Recognize Healthy Body System 9yESHEy T100% (2)
- Endocrine System: Definition & BackgroundDocument11 pagesEndocrine System: Definition & Backgroundعبدالرحمن عدي عبدالفتاحNo ratings yet
- Anatomy of The Endocrine Glands: HypothalamusDocument10 pagesAnatomy of The Endocrine Glands: Hypothalamusgashbin latifNo ratings yet
- Histology Lec-11 EndocrineDocument10 pagesHistology Lec-11 EndocrineKevin C. AguilarNo ratings yet
- Describe and Explained Organs Belong To Endocrine SystemDocument4 pagesDescribe and Explained Organs Belong To Endocrine SystemnilaahanifahNo ratings yet
- Saq 2 EmbryologyDocument8 pagesSaq 2 EmbryologyShilpa DuttaNo ratings yet
- Endo All MergeDocument586 pagesEndo All MergeShivani DurgeNo ratings yet
- Lab Act 3 HistologyDocument15 pagesLab Act 3 HistologydiithandariNo ratings yet
- PPP Embryo N Histo Endocrine SystemDocument32 pagesPPP Embryo N Histo Endocrine SystemMarisol AcostaNo ratings yet
- BiologyDocument21 pagesBiologynitesh.th15005No ratings yet
- Embryology and Histology of Endocrine Glands: Dr. Ramada Khasawneh 2020Document61 pagesEmbryology and Histology of Endocrine Glands: Dr. Ramada Khasawneh 2020Qutaybah JahmanyNo ratings yet
- Endocrine System (Finals)Document36 pagesEndocrine System (Finals)hjygcaingodNo ratings yet
- C10 Endocrine SystemDocument24 pagesC10 Endocrine Systemwajomi100% (1)
- Disorders of The Nervous System2Document20 pagesDisorders of The Nervous System2Ian Rizavi Villamor AntopinaNo ratings yet
- 03a Nervous System in FishesDocument7 pages03a Nervous System in FishesYudi ArtNo ratings yet
- Body System II 7 Endo 1Document27 pagesBody System II 7 Endo 1rrq8cwk2gnNo ratings yet
- Endocrine GlandsDocument24 pagesEndocrine GlandsZoyaNo ratings yet
- Erm BH E4w1Document7 pagesErm BH E4w1Emiliano RiquelmeNo ratings yet
- W4-15 Endocrine and Alimentary Layers of The Neck Lecture PDFDocument75 pagesW4-15 Endocrine and Alimentary Layers of The Neck Lecture PDFSAHIL AGARWALNo ratings yet
- Endocrine SysDocument17 pagesEndocrine SyssjkNo ratings yet
- Endocrinology 2Document13 pagesEndocrinology 2Dr.Mahesh kumar GuptaNo ratings yet
- Anatomy of Nervous SystemDocument59 pagesAnatomy of Nervous SystemSeema ChapagainNo ratings yet
- Endocrine System: Anatomy 1 Stage - College of PharmacyDocument6 pagesEndocrine System: Anatomy 1 Stage - College of PharmacyMarci MunirNo ratings yet
- Science 6 - Week 4 - 2nd QuarterDocument5 pagesScience 6 - Week 4 - 2nd QuarterHoneylet Alberto Castro ArilloNo ratings yet
- Histology of Endocrine SystemDocument35 pagesHistology of Endocrine SystemX And ZNo ratings yet
- Hesto 1Document46 pagesHesto 1zsf8m52ky4No ratings yet
- Physio RespiDocument17 pagesPhysio RespiJenica SorianoNo ratings yet
- Downloadfile 4Document49 pagesDownloadfile 4shuvo devNo ratings yet
- Endocrine SystemDocument6 pagesEndocrine Systemrk749vbsk6No ratings yet
- Class - XI - M 3 PPT XI Bio CH 7 STRUCTURAL ORGANISATION IN ANIMALSCockroachDocument23 pagesClass - XI - M 3 PPT XI Bio CH 7 STRUCTURAL ORGANISATION IN ANIMALSCockroachkaran.krisn1No ratings yet
- Nervous System of The FrogDocument38 pagesNervous System of The Frognone57% (7)
- Anaphy LabDocument5 pagesAnaphy Labmariajepoi101210No ratings yet
- Epithelial Tissue Stations 1-3Document4 pagesEpithelial Tissue Stations 1-3api-692335939No ratings yet
- Pituitary GlandDocument66 pagesPituitary GlandAsmita BhattNo ratings yet
- The Somatic Nervous System (SNS or Voluntary Nervous System) Is The Part of The Associated With The Voluntary Control of Body Movements ViaDocument3 pagesThe Somatic Nervous System (SNS or Voluntary Nervous System) Is The Part of The Associated With The Voluntary Control of Body Movements ViaAkash SahaNo ratings yet
- HomeostasisDocument24 pagesHomeostasismark angelesNo ratings yet
- The BrainDocument103 pagesThe Brainrenzvalorant28No ratings yet
- Endocrine Gland....Document62 pagesEndocrine Gland....Sabita TripathiNo ratings yet
- Rat Dissection Student VersionDocument11 pagesRat Dissection Student VersionKong Hung Xi100% (1)
- WWW Kenhub Com en Library Anatomy Endocrine SystemDocument20 pagesWWW Kenhub Com en Library Anatomy Endocrine SystemIvana Odelia NainggolanNo ratings yet
- 12 - Endocrine SystemDocument20 pages12 - Endocrine SystemNovie Roycell Fernandez RueloNo ratings yet
- المحاضرة السادسة Chordata المختصرةDocument28 pagesالمحاضرة السادسة Chordata المختصرةAhmed OrabyNo ratings yet
- Disorders of The Nervous SystemDocument26 pagesDisorders of The Nervous Systemshishiminaj100% (1)
- Gen BioDocument11 pagesGen BioIrish MaireignNo ratings yet
- 1.1 Community Engagement Solidarity and Citizenship CSC - Compendium of Appendices For DLPs - Class FDocument11 pages1.1 Community Engagement Solidarity and Citizenship CSC - Compendium of Appendices For DLPs - Class Fherlyn jueloNo ratings yet
- Endocrine SystemDocument25 pagesEndocrine SystemLits KhoNo ratings yet
- Rat Dissection Lab UpdatedDocument8 pagesRat Dissection Lab UpdatedStephanie LeeNo ratings yet
- LP 17 MG Engl - EndocrineDocument3 pagesLP 17 MG Engl - EndocrinedianaNo ratings yet
- Chapter 2. Structure and Functions of Important Endocrine GlandsDocument17 pagesChapter 2. Structure and Functions of Important Endocrine GlandsAmit SharmaNo ratings yet
- Muslimah D. Diangca - Activity 4Document4 pagesMuslimah D. Diangca - Activity 4Baby Corner MarawiNo ratings yet
- Zodiac Academy: Hari Krishna G L Academic CoordinatorDocument94 pagesZodiac Academy: Hari Krishna G L Academic CoordinatorHARI KRISHNA G LNo ratings yet
- Endocrine System Group 9Document77 pagesEndocrine System Group 9Jei SanNo ratings yet
- HypothalamusDocument39 pagesHypothalamusKhalid AlhemyariNo ratings yet
- AD 7 The Nervous SystemDocument28 pagesAD 7 The Nervous SystemBernard SalimNo ratings yet
- BZ Lab 2Document7 pagesBZ Lab 2Alexa Jean D. HonrejasNo ratings yet
- The 96Document10 pagesThe 96Andrei AlvaradoNo ratings yet
- Thyroid: Lamina Pretrachealis Fasciae CervicalisDocument5 pagesThyroid: Lamina Pretrachealis Fasciae CervicalisbhaveshrockzzNo ratings yet
- Syaraf Indera PDFDocument61 pagesSyaraf Indera PDFWisdanuar Yudhasmara WijanarkoNo ratings yet
- Summary of Stanley Rosenberg's Accessing the Healing Power of the Vagus NerveFrom EverandSummary of Stanley Rosenberg's Accessing the Healing Power of the Vagus NerveNo ratings yet
- Saida Hal 8-18 "Analisis Faktor Risiko Kejadian Hipertensi ... " Jurnal KeperawatanDocument11 pagesSaida Hal 8-18 "Analisis Faktor Risiko Kejadian Hipertensi ... " Jurnal KeperawatanYanti HarjonoNo ratings yet
- PMLS Midterm ReviewerDocument41 pagesPMLS Midterm ReviewerGwen YosheenNo ratings yet
- SONI - Fundamentals of Botany - Vol 2 (2011, MC GRAW HILL INDIA) PDFDocument398 pagesSONI - Fundamentals of Botany - Vol 2 (2011, MC GRAW HILL INDIA) PDFPrince Quokka Rai100% (5)
- Collagen ProteoglycansDocument2 pagesCollagen ProteoglycansAlliah CasingNo ratings yet
- Blood Questions: (Total 5 Marks)Document11 pagesBlood Questions: (Total 5 Marks)T-Jay Ellis-DaleNo ratings yet
- Drugs Acting On The Renin-Angiotensin-Aldosterone SystemDocument2 pagesDrugs Acting On The Renin-Angiotensin-Aldosterone SystemKate EvangelistaNo ratings yet
- Special Procedures Q and ADocument11 pagesSpecial Procedures Q and Ajustine gailNo ratings yet
- As-Level Biology Cell Membranes and Transport Worksheet: Name: - Class: XI SCIENCEDocument5 pagesAs-Level Biology Cell Membranes and Transport Worksheet: Name: - Class: XI SCIENCEDina Anggraini Pramitasari100% (1)
- LST RO3 ManuscriptDocument76 pagesLST RO3 Manuscriptjoshua bravoNo ratings yet
- Biology Syllabus GR11-12 (2022-2023)Document37 pagesBiology Syllabus GR11-12 (2022-2023)Amal HassanNo ratings yet
- LED Wavelenghts and Intensities in Cherry RootstockDocument25 pagesLED Wavelenghts and Intensities in Cherry RootstockVinicius BohryNo ratings yet
- Class 2 Lesson PlanDocument16 pagesClass 2 Lesson PlanArjun Neupane100% (1)
- Scoliosis: Causes, Genetics, Symptoms, and Treatment in A Southern Italy PopulationDocument48 pagesScoliosis: Causes, Genetics, Symptoms, and Treatment in A Southern Italy PopulationKirolos WassefNo ratings yet
- General Biology 2nd Quarter ReviewerDocument14 pagesGeneral Biology 2nd Quarter ReviewerJaime TorresNo ratings yet
- Diagnosis and Medical Management of Parkinson.5Document20 pagesDiagnosis and Medical Management of Parkinson.5Gustavo Silva SchafascheckNo ratings yet
- Mark Scheme (Results) Summer 2015: Pearson Edexcel International GCSE in Human Biology (4HB0) Paper 02Document14 pagesMark Scheme (Results) Summer 2015: Pearson Edexcel International GCSE in Human Biology (4HB0) Paper 02Joseph LAU [11D]No ratings yet
- KS3 Science Workbook June July PDFDocument56 pagesKS3 Science Workbook June July PDFManimekalaiNo ratings yet
- First Year LMD Phonetics Full GuideDocument106 pagesFirst Year LMD Phonetics Full GuideGrX4 ChemsoNo ratings yet
- Nephrotic SyndromeDocument38 pagesNephrotic SyndromeSwati SharmaNo ratings yet
- Functions of The Human Respiratory System: What's inDocument4 pagesFunctions of The Human Respiratory System: What's injoyNo ratings yet
- General Biology 2: Quarter 4: Week 2 - Module 1B PDocument25 pagesGeneral Biology 2: Quarter 4: Week 2 - Module 1B PSyrine Myles Sullivan100% (2)
- Forensic Medicine: It Is The Sum Total of All Reaction of Tissue or Organ To TraumaDocument2 pagesForensic Medicine: It Is The Sum Total of All Reaction of Tissue or Organ To TraumaLex Tamen CoercitorNo ratings yet
- Introduction To UrinalysisDocument8 pagesIntroduction To UrinalysisKyle PicocNo ratings yet
- Biochemical Analysis of BloodDocument2 pagesBiochemical Analysis of Bloodbucsit100% (1)
- Chapter 11Document4 pagesChapter 11Nona PeriarceNo ratings yet
- 9700 - s20 - Ms - 42 Biologya2Document20 pages9700 - s20 - Ms - 42 Biologya2Fatahna KhanzadaNo ratings yet
- Biology SL P2Document8 pagesBiology SL P2KenanNo ratings yet
- 1 - Anatomical Terminology KEYDocument6 pages1 - Anatomical Terminology KEY愛.NiniNo ratings yet
- Class 9 MT Science Notes TissueDocument12 pagesClass 9 MT Science Notes TissueKhushi Agrawal100% (1)
- 4HB0 01 Rms 2018030Document16 pages4HB0 01 Rms 2018030hasan memosNo ratings yet