You might also like
- ReplikacijaDocument54 pagesReplikacijabetmenNo ratings yet
- GENETIKADocument8 pagesGENETIKANikol RadovićNo ratings yet
- Replikacija, Održavanje I Preslagivanje Genomske DNA: Molekularna Biologija Poglavlje 6Document19 pagesReplikacija, Održavanje I Preslagivanje Genomske DNA: Molekularna Biologija Poglavlje 6Shingeki No ObamaNo ratings yet
- Genetika, 1.kolokvij, Prva VerzijaDocument7 pagesGenetika, 1.kolokvij, Prva VerzijaIvan PetrikNo ratings yet
- GenDocument45 pagesGenPetra VincekovićNo ratings yet
- Bio IspitDocument2 pagesBio IspitTin GvozdanovicNo ratings yet
- Biologija Sa Humanom Genetikom - Vjezba 3Document38 pagesBiologija Sa Humanom Genetikom - Vjezba 3Osman PobrićNo ratings yet
- Biologija DM Skripta Jesen2014 3Document63 pagesBiologija DM Skripta Jesen2014 3Kristina BukovacNo ratings yet
- Vježba Br. 2 Molekularna BiologijaDocument14 pagesVježba Br. 2 Molekularna BiologijaBosna IslamNo ratings yet
- Replikacija SkracenoDocument4 pagesReplikacija Skracenojakov12345No ratings yet
- Molekularna Biologija 2Document33 pagesMolekularna Biologija 2Ismar Mujagic100% (1)
- Jezgra I Jezgrica PrezentacijaDocument56 pagesJezgra I Jezgrica PrezentacijaIvana05100% (1)
- 4 Poglavlje - Staniƒni GenomiDocument35 pages4 Poglavlje - Staniƒni GenomiAdnanHusicNo ratings yet
- Gen 2Document59 pagesGen 2a_delNo ratings yet
- Uvod U Molekularnu BiologijuDocument36 pagesUvod U Molekularnu Biologijumaestro95100% (1)
- Genetika I.dioDocument49 pagesGenetika I.dioMatiA MMNo ratings yet
- Dna U BrojkamaDocument31 pagesDna U BrojkamaMihael DelićNo ratings yet
- METODEDocument20 pagesMETODEtisak.kandijaNo ratings yet
- BIOLOGIJADocument3 pagesBIOLOGIJAKarmenNo ratings yet
- RNA I DNA. Tijek Genetičke InformacijeDocument8 pagesRNA I DNA. Tijek Genetičke InformacijeJelena KrstanovićNo ratings yet
- Jedro - VIII Poglavlje Stanica-CooperDocument26 pagesJedro - VIII Poglavlje Stanica-CooperEmina OmeragićNo ratings yet
- 5 Poglavlje - Replikacija DNADocument33 pages5 Poglavlje - Replikacija DNAAdnanHusicNo ratings yet
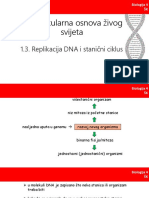
- Molekularna Osnova Živog Svijeta-Replikacija DNA I Stanični CiklusDocument42 pagesMolekularna Osnova Živog Svijeta-Replikacija DNA I Stanični Ciklusnea100% (1)
- Udvostručenje DNADocument2 pagesUdvostručenje DNAsuziNo ratings yet
- Popravak DNKDocument10 pagesPopravak DNKMiroslav TrickovicNo ratings yet
- JezgraDocument48 pagesJezgrafernandarubiNo ratings yet
- Središnja Dogma Molekularne BiologijeDocument1 pageSredišnja Dogma Molekularne BiologijeKarlo KoširNo ratings yet
- BiologijaDocument10 pagesBiologijaTamata BeberNo ratings yet
- Sekvenciranje DNADocument20 pagesSekvenciranje DNAsiranzikaNo ratings yet
- Ishodi - MolBioDocument79 pagesIshodi - MolBioLucija PavkovicNo ratings yet
- Skripta BiologijaDocument29 pagesSkripta BiologijaAdmir AbdihodzicNo ratings yet
- GenetikaDocument17 pagesGenetikadositejmNo ratings yet
- BIOLOGIJADocument233 pagesBIOLOGIJAIvica Markić100% (1)
- Biolog I JaaaDocument6 pagesBiolog I JaaaEnes KubatNo ratings yet
- Replikacija DnaDocument1 pageReplikacija DnaDominik OdmorekNo ratings yet
- Kemijska Osnova NasljeđivanjaDocument1 pageKemijska Osnova NasljeđivanjaKarlo KoširNo ratings yet
- Gene TikaDocument33 pagesGene TikadubravkagvozdicNo ratings yet
- Maturalni Rad Kromosomske AberacijeDocument25 pagesMaturalni Rad Kromosomske Aberacijecika_mraz67% (3)
- ReplikacijaDocument25 pagesReplikacijatema_racamaNo ratings yet
- Sekvenciranje DNKDocument24 pagesSekvenciranje DNKSanja ZukićNo ratings yet
- Genetika Skripta Za Dodatnu Nastavu Popravljeno PDFDocument168 pagesGenetika Skripta Za Dodatnu Nastavu Popravljeno PDFmikiksenija50% (2)
- Struktura I Funkcija Neurona HANDOUT Osnove Biološke PsihologijeDocument5 pagesStruktura I Funkcija Neurona HANDOUT Osnove Biološke PsihologijeLana OstojićNo ratings yet
- 9.nastavni Tekst Na webu-SKRIPTA IZ KOLEGIJA Osnove Fiziologije CovjekaDocument99 pages9.nastavni Tekst Na webu-SKRIPTA IZ KOLEGIJA Osnove Fiziologije CovjekaMartina MajerNo ratings yet
- PCR KnjigaDocument6 pagesPCR KnjigaAnel Ward RedzicNo ratings yet
- 6. Molekula DNADocument11 pages6. Molekula DNALara ZivkovicNo ratings yet
- Medicinska Biologija Ishodi UcenjaDocument12 pagesMedicinska Biologija Ishodi UcenjaLovro DruzijanicNo ratings yet
- Stanični Ciklus Eukariota, MitozaDocument7 pagesStanični Ciklus Eukariota, Mitozasana4nnaNo ratings yet
- Prokarioti - MejozaDocument5 pagesProkarioti - MejozaMateo BenčekNo ratings yet
- Ćelijska DiobaDocument11 pagesĆelijska DiobaDavor TunjićNo ratings yet
- Biologija Sa Humanom GenetikomDocument12 pagesBiologija Sa Humanom GenetikomVedrana FrancuzNo ratings yet
- Strateški MenadžmentDocument17 pagesStrateški MenadžmentSalih KavazovicNo ratings yet
- Strateški MenadžmentDocument17 pagesStrateški MenadžmentSalih KavazovicNo ratings yet
- Strateški MenadžmentDocument17 pagesStrateški MenadžmentSalih KavazovicNo ratings yet
- Strateški MenadžmentDocument17 pagesStrateški MenadžmentSalih KavazovicNo ratings yet
- Pravilnik o Zavrsnom Radu Prvog Ciklusa StudijaDocument9 pagesPravilnik o Zavrsnom Radu Prvog Ciklusa StudijaSalih KavazovicNo ratings yet
- Ebay Marketing MixDocument3 pagesEbay Marketing MixSalih KavazovicNo ratings yet
- Zadaća Iz ViMPDocument9 pagesZadaća Iz ViMPSalih KavazovicNo ratings yet
- Zadaća Iz ViMPDocument9 pagesZadaća Iz ViMPSalih KavazovicNo ratings yet
- Zadaća Iz ViMPDocument9 pagesZadaća Iz ViMPSalih KavazovicNo ratings yet
- Strateški MenadžmentDocument17 pagesStrateški MenadžmentSalih KavazovicNo ratings yet
- Mva Pitanja I OdgovoriDocument6 pagesMva Pitanja I OdgovoriSalih KavazovicNo ratings yet
- Zadaća Iz ViMPDocument9 pagesZadaća Iz ViMPSalih KavazovicNo ratings yet
- Mva Pitanja I OdgovoriDocument6 pagesMva Pitanja I OdgovoriSalih KavazovicNo ratings yet
- Zina OkićDocument1 pageZina OkićSalih KavazovicNo ratings yet
- Mva Pitanja I OdgovoriDocument6 pagesMva Pitanja I OdgovoriSalih KavazovicNo ratings yet
- Zux-4 LetakDocument6 pagesZux-4 LetakSalih KavazovicNo ratings yet
- O NAMA StaroDocument2 pagesO NAMA StaroSalih KavazovicNo ratings yet
- Popravak DNA SeminarskiDocument19 pagesPopravak DNA SeminarskiSalih KavazovicNo ratings yet
- Replikacija DNK 1Document2 pagesReplikacija DNK 1Salih KavazovicNo ratings yet
- Puska 1Document3 pagesPuska 1Salih KavazovicNo ratings yet
- Zina OkićDocument1 pageZina OkićSalih KavazovicNo ratings yet
- Zux 4Document2 pagesZux 4Salih KavazovicNo ratings yet