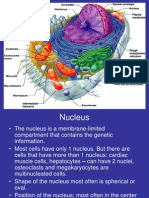
You might also like
- Wound Healing SchwartzDocument37 pagesWound Healing SchwartzCarla Putri Chandra100% (1)
- Vox CorporisDocument898 pagesVox CorporisSushiNo ratings yet
- Text Book Questions and Answers Biology 30Document44 pagesText Book Questions and Answers Biology 30khristina_holmesNo ratings yet
- IB Biology Study NotesDocument134 pagesIB Biology Study Noteshdpham100% (25)
- Lesson-Plan-in-Science-8 - Cell DivisionDocument18 pagesLesson-Plan-in-Science-8 - Cell DivisionMaricris Ranara100% (1)
- Pathophysiology of PneumoniaDocument4 pagesPathophysiology of PneumoniaShermane Criszen F. Sallan100% (4)
- The Pulp 1 SummaryDocument46 pagesThe Pulp 1 SummaryVassili DimogluNo ratings yet
- Lab 9Document40 pagesLab 9bob0% (1)
- The Cell Cycle Activity 1Document5 pagesThe Cell Cycle Activity 1sugarxglossNo ratings yet
- The Nucleus and Nuclear EnvelopeDocument58 pagesThe Nucleus and Nuclear EnvelopeImperiall100% (1)
- Cell DivisionDocument31 pagesCell DivisionShin Thant Ko B10 June SectionNo ratings yet
- Cell Division: By: Grace Nichols, Coral Friend, and Amanda Trussell Mr. Taylor's 8 Grade Gifted Class January, 2006Document17 pagesCell Division: By: Grace Nichols, Coral Friend, and Amanda Trussell Mr. Taylor's 8 Grade Gifted Class January, 2006Rahul UniyalNo ratings yet
- Cell Cycle and Cell DivisionDocument13 pagesCell Cycle and Cell DivisionHeerak SahuNo ratings yet
- Chapter Ten:: Cell Cycle and Cell DivisionDocument12 pagesChapter Ten:: Cell Cycle and Cell DivisionShivani Shree SundaramoorthyNo ratings yet
- Cell Biology Handout - 1Document62 pagesCell Biology Handout - 1Dewi Lasimpara100% (1)
- Cell Division - 2.structure of Chromosomes - 3.principles of GeneticsDocument148 pagesCell Division - 2.structure of Chromosomes - 3.principles of Geneticsankurbiology0% (1)
- Chapter 10 Cell Cycle and Cell DivisionDocument14 pagesChapter 10 Cell Cycle and Cell DivisionPhoenix RockiNo ratings yet
- Biology Unit 16Document14 pagesBiology Unit 16joyNo ratings yet
- How Bacteria and Eukaryotes Divide CellsDocument69 pagesHow Bacteria and Eukaryotes Divide CellsAhmed MohamadNo ratings yet
- Mitosis: Not To Be Confused With, ,, orDocument6 pagesMitosis: Not To Be Confused With, ,, orWacek4444No ratings yet
- Cell CycleDocument111 pagesCell Cyclepreyaaa pascuaNo ratings yet
- Ang Siklo NG SikhayDocument71 pagesAng Siklo NG SikhayJohn Lenard BerbiscoNo ratings yet
- Lesson 4Document8 pagesLesson 4Marie Antonette BaligodNo ratings yet
- Cell Cycle Stages and Phases ExplainedDocument43 pagesCell Cycle Stages and Phases ExplainedMariam QaisNo ratings yet
- Allot Mindorff Mitosis and MeiosisDocument11 pagesAllot Mindorff Mitosis and MeiosisElTrucoCeroNo ratings yet
- 0 BooksDocument20 pages0 Bookslaraib zahidNo ratings yet
- Cell Cycle Part 1 (Mitosis)Document25 pagesCell Cycle Part 1 (Mitosis)Awais GulNo ratings yet
- Presentation on the Cleavage ProcessDocument28 pagesPresentation on the Cleavage ProcessAkpene RachealNo ratings yet
- Mitosis Lab ProceduresDocument16 pagesMitosis Lab ProceduresDavid AhneNo ratings yet
- Modules 1 Cells Its PhysiologyDocument6 pagesModules 1 Cells Its PhysiologySheena Marie M. TarleNo ratings yet
- Army Public School Gopalpur: Class - 11 Science Subject - BiologyDocument6 pagesArmy Public School Gopalpur: Class - 11 Science Subject - BiologyAshok KumarNo ratings yet
- Plant Cell E-BookDocument21 pagesPlant Cell E-BookSammy ShnayderNo ratings yet
- Cell DivisionDocument24 pagesCell DivisionSam MumoNo ratings yet
- Bio 1Document6 pagesBio 1zachharveyneilNo ratings yet
- Mitosis Thesis StatementDocument7 pagesMitosis Thesis Statementzidhelvcf100% (2)
- GENERAL BIOLOGYDocument5 pagesGENERAL BIOLOGYFern Nicole TagatacNo ratings yet
- Cell Cycle and Mitosis Grade 7 BiologyDocument22 pagesCell Cycle and Mitosis Grade 7 BiologyLaszlo ArdaiNo ratings yet
- FSC111 - Cell Structure and FunctionDocument15 pagesFSC111 - Cell Structure and FunctionKenneth SilvaNo ratings yet
- Cell Cycle: What Is It? Why Do We Nee D It?Document28 pagesCell Cycle: What Is It? Why Do We Nee D It?suvendumitraNo ratings yet
- Week 7 Mitosis and GeneticsDocument32 pagesWeek 7 Mitosis and GeneticsMierja Van ZuraNo ratings yet
- BIOL223-Lab 7Document40 pagesBIOL223-Lab 7chicken fries100% (2)
- Baghdad College of Medical Science (Human Biology)Document9 pagesBaghdad College of Medical Science (Human Biology)علي صلاح مهديNo ratings yet
- Mitotic Cell Division - Exercise 7: ObjectivesDocument56 pagesMitotic Cell Division - Exercise 7: Objectiveskaran79No ratings yet
- Assignment 01, Week 2Document11 pagesAssignment 01, Week 2padhiyararpitaNo ratings yet
- 3 - Mitosis & Meiosis Voice OverDocument11 pages3 - Mitosis & Meiosis Voice OverOthman AlharounNo ratings yet
- Cell Cycle: The Stages of Growth and DivisionDocument20 pagesCell Cycle: The Stages of Growth and DivisionUzma AdnanNo ratings yet
- I. Embryology: Yolk.-The Yolk Comprises (1) The Cytoplasm of The Ordinary Animal Cell With Its Spongioplasm andDocument7 pagesI. Embryology: Yolk.-The Yolk Comprises (1) The Cytoplasm of The Ordinary Animal Cell With Its Spongioplasm andEzra GenerosoNo ratings yet
- I. Embryology: Yolk.-The Yolk Comprises (1) The Cytoplasm of The Ordinary Animal Cell With Its Spongioplasm andDocument7 pagesI. Embryology: Yolk.-The Yolk Comprises (1) The Cytoplasm of The Ordinary Animal Cell With Its Spongioplasm andEzra GenerosoNo ratings yet
- Bio NotesDocument6 pagesBio NotesHuck KimNo ratings yet
- Bio HomeworkDocument1 pageBio HomeworkshamooNo ratings yet
- Whole Cell ActivityDocument16 pagesWhole Cell ActivityEugene VinaraoNo ratings yet
- Chapter 06Document17 pagesChapter 06homamunfatNo ratings yet
- Cell DivisionDocument10 pagesCell DivisionVardan ShrivastavaNo ratings yet
- Cell CycleDocument9 pagesCell CycleCrystal Joy RebelloNo ratings yet
- Cell Division and Reproduction ExplainedDocument29 pagesCell Division and Reproduction ExplainedEdel John SivaNo ratings yet
- BIO111 NEW1 CompleteDocument25 pagesBIO111 NEW1 Completealiu sodiqNo ratings yet
- THE CELL II: Nucleus, Cell Cycle, Cell Division, ApoptosisDocument23 pagesTHE CELL II: Nucleus, Cell Cycle, Cell Division, ApoptosisDebby14No ratings yet
- Cell Cycle MitosisDocument39 pagesCell Cycle MitosisGianne Louise SilvestreNo ratings yet
- AP Bio Lab 3 FinalDocument6 pagesAP Bio Lab 3 Finalniteeshsettypalli100% (1)
- רזי 7 חלוקת תאDocument24 pagesרזי 7 חלוקת תאBoba BanayNo ratings yet
- Cell Cycle & MitosisDocument42 pagesCell Cycle & Mitosismariam yahiaNo ratings yet
- Section 3 - The Mitotic Cycle 1Document36 pagesSection 3 - The Mitotic Cycle 1abielgtgNo ratings yet
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Every Child Matters: Change For Children: in Social CareDocument16 pagesEvery Child Matters: Change For Children: in Social CarehomamunfatNo ratings yet
- GCSE ENG REVISED Past Papers Mark Schemes Standard January Series 2014 13606Document11 pagesGCSE ENG REVISED Past Papers Mark Schemes Standard January Series 2014 13606homamunfatNo ratings yet
- GCSE ENG REVISED Past Papers Mark Schemes Standard January Series 2014 13603Document13 pagesGCSE ENG REVISED Past Papers Mark Schemes Standard January Series 2014 13603homamunfatNo ratings yet
- GCSE ENG REVISED Past Papers Mark Schemes Standard MayJune Series 2011 8651Document8 pagesGCSE ENG REVISED Past Papers Mark Schemes Standard MayJune Series 2011 8651homamunfatNo ratings yet
- GCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9985Document16 pagesGCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9985homamunfatNo ratings yet
- 50+ ActivitiiesDocument56 pages50+ ActivitiieshomamunfatNo ratings yet
- Gcse Eng Revised Support 6628Document79 pagesGcse Eng Revised Support 6628homamunfat100% (1)
- GCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9984Document12 pagesGCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9984homamunfatNo ratings yet
- GCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9983Document16 pagesGCSE ELIT REVISED Past Papers Mark Schemes Standard MayJune Series 2011 9983homamunfatNo ratings yet
- 18 Yatcb Lesson Plans Complete Book FINALDocument146 pages18 Yatcb Lesson Plans Complete Book FINALhomamunfatNo ratings yet
- 50 Word Story TellingDocument1 page50 Word Story TellinghomamunfatNo ratings yet
- B. Red, Yellow, and Blue Can Be Mixed Together To Make Secondary ColorsDocument1 pageB. Red, Yellow, and Blue Can Be Mixed Together To Make Secondary ColorshomamunfatNo ratings yet
- Colors: Lesson Ten: The Three Primary Colors Are Red, Yellow, and Blue. They CanDocument1 pageColors: Lesson Ten: The Three Primary Colors Are Red, Yellow, and Blue. They CanhomamunfatNo ratings yet
- 23.2 Writing An A - Essay - A ModelDocument2 pages23.2 Writing An A - Essay - A ModelhomamunfatNo ratings yet
- 23.1 Writing A Band 5 EssayDocument5 pages23.1 Writing A Band 5 EssayhomamunfatNo ratings yet
- 23.1 Writing A Band 5 EssayDocument5 pages23.1 Writing A Band 5 EssayhomamunfatNo ratings yet
- 23.1 Writing A Band 5 EssayDocument5 pages23.1 Writing A Band 5 EssayhomamunfatNo ratings yet
- 13.05.15 Prepositions of PlaceDocument1 page13.05.15 Prepositions of PlacehomamunfatNo ratings yet
- GCSE English Language Unit 3, The Crucible by Arthur MillerDocument2 pagesGCSE English Language Unit 3, The Crucible by Arthur MillerhomamunfatNo ratings yet
- 15.1 Speaking and Listening Assessment - Role PlayingDocument2 pages15.1 Speaking and Listening Assessment - Role PlayinghomamunfatNo ratings yet
- 16Document29 pages16homamunfatNo ratings yet
- GCSE English Language Unit 3, The Crucible by Arthur MillerDocument2 pagesGCSE English Language Unit 3, The Crucible by Arthur MillerhomamunfatNo ratings yet
- 13.05.15 Street Map For ComprehensionDocument2 pages13.05.15 Street Map For ComprehensionhomamunfatNo ratings yet
- 13.05.15 Street Map Graded QuestionsDocument2 pages13.05.15 Street Map Graded QuestionshomamunfatNo ratings yet
- Learn spatial words like between and oppositeDocument1 pageLearn spatial words like between and oppositehomamunfatNo ratings yet
- 13.05.15 Blank Map InstructionsDocument1 page13.05.15 Blank Map InstructionshomamunfatNo ratings yet
- 13.05.15 Street Map For ComprehensionDocument2 pages13.05.15 Street Map For ComprehensionhomamunfatNo ratings yet
- B. Write The Following Information On The Calendar On Page 10. Write SmallDocument1 pageB. Write The Following Information On The Calendar On Page 10. Write SmallhomamunfatNo ratings yet
- C. Write The Missing Days and Questions in The Boxes. Start The Questions WithDocument1 pageC. Write The Missing Days and Questions in The Boxes. Start The Questions WithVero NikaNo ratings yet
- 11 CalendarsDocument1 page11 CalendarsIoana TralalaNo ratings yet
- Muscle Structure and Meat CompositionDocument18 pagesMuscle Structure and Meat CompositionJoy April Aguilar DeGuzmanNo ratings yet
- BiologyDocument11 pagesBiologykpop trashNo ratings yet
- Gell & Coombs Classification of Hypersensitivity ReactionDocument17 pagesGell & Coombs Classification of Hypersensitivity ReactionShia LevyNo ratings yet
- Plasma Proteins - PPT 93Document27 pagesPlasma Proteins - PPT 93Dr.P.NatarajanNo ratings yet
- PhysioEx Exercise 3 Activity 1Document8 pagesPhysioEx Exercise 3 Activity 1tasyaauliatabina1122No ratings yet
- Quick Relaxation StrategiesDocument2 pagesQuick Relaxation Strategiesaahlada srividyaNo ratings yet
- The Cosmic HealingDocument8 pagesThe Cosmic Healingvince talzNo ratings yet
- Test DirectoryDocument163 pagesTest DirectoryHooshmandKaafNo ratings yet
- Pedh 112 Week 11 19Document11 pagesPedh 112 Week 11 19April Puncia67% (3)
- Gaucher Disease PDFDocument3 pagesGaucher Disease PDFKatelyn ColleyNo ratings yet
- Purtscher Retinopathy EyenetDocument4 pagesPurtscher Retinopathy EyenetMila KurniasariNo ratings yet
- Thesis by Woong Kim SmallerDocument415 pagesThesis by Woong Kim SmallerWoong KimNo ratings yet
- Suctioning Techniques and ProceduresDocument24 pagesSuctioning Techniques and Proceduresluis_chubee100% (1)
- Optimal Breathing® Mastery CourseDocument74 pagesOptimal Breathing® Mastery Coursecreat1100% (1)
- Pcol Prelim-LjmDocument10 pagesPcol Prelim-LjmLyka MarceloNo ratings yet
- Jntu Kakinada 3rd B.pharm 1st Semester Syllabus r10Document17 pagesJntu Kakinada 3rd B.pharm 1st Semester Syllabus r10vinay0717No ratings yet
- Project 2 ExampleDocument5 pagesProject 2 ExampleCzar ParrenoNo ratings yet
- Fisilogi Anal KanalDocument45 pagesFisilogi Anal KanalMargarethaNo ratings yet
- Cardiac CycleDocument4 pagesCardiac CyclefailinNo ratings yet
- Prepared By: Ruby Angela D. O. PenaDocument22 pagesPrepared By: Ruby Angela D. O. PenaFunnish KumarNo ratings yet
- Chapter 7 Section 2 OutlineDocument5 pagesChapter 7 Section 2 Outlineapi-263455051100% (1)
- Hyperemesis GravidarumDocument16 pagesHyperemesis GravidarumBabi PanggangNo ratings yet
- 2020 Immediate Care of The Newborn 3Document56 pages2020 Immediate Care of The Newborn 3Ellah PerenioNo ratings yet
- Mechanical Ventilation and Intracranial PressureDocument30 pagesMechanical Ventilation and Intracranial PressureFlavius AnghelNo ratings yet
- Agha Rahim - Defense AttachmentDocument30 pagesAgha Rahim - Defense AttachmentLuis HernandezNo ratings yet
- Roots Prefixes and Suffixes in BiologyDocument9 pagesRoots Prefixes and Suffixes in Biologyapi-240662720No ratings yet