You might also like
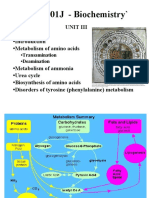
- Lesson 20 General Pathways of Amino Acids Transformation, Transamination, and Decarboxylation. The Role of Enzymes and Coenzymes.Document87 pagesLesson 20 General Pathways of Amino Acids Transformation, Transamination, and Decarboxylation. The Role of Enzymes and Coenzymes.Mi PatelNo ratings yet
- Questions On Amino Acid Urea CycleDocument17 pagesQuestions On Amino Acid Urea Cyclejmenchaca8080No ratings yet
- 9a. Metabolisme Asam AminoDocument62 pages9a. Metabolisme Asam AminohimawarumNo ratings yet
- Screenshot 2021-05-25 at 1.47.14 PMDocument7 pagesScreenshot 2021-05-25 at 1.47.14 PMAynaz ChanNo ratings yet
- Renal Summary 1.21 On Urea Production, Amino Acid Synthesis and MetabolismDocument4 pagesRenal Summary 1.21 On Urea Production, Amino Acid Synthesis and Metabolismvivienne_fokNo ratings yet
- Removal of Nitrogen From Amino AcidsDocument34 pagesRemoval of Nitrogen From Amino AcidsSafura IjazNo ratings yet
- 2023 2024 b3502 Lecture 18 Protein MetabolismDocument21 pages2023 2024 b3502 Lecture 18 Protein Metabolismpitsolerato60No ratings yet
- Metabolisme ProteinDocument64 pagesMetabolisme ProteinErlin GafrinNo ratings yet
- Class 3rdDocument16 pagesClass 3rdJude Chisom Jnr NwaugoNo ratings yet
- METABOLISME ASAM AMINO Protein BiologiDocument31 pagesMETABOLISME ASAM AMINO Protein BiologiAxzchiuu :vNo ratings yet
- Amino Acid CatabolismDocument7 pagesAmino Acid CatabolismAhmed Ali Mohammed AlbashirNo ratings yet
- Item - 11, Protein MetabolismDocument12 pagesItem - 11, Protein MetabolismSheikh FahadNo ratings yet
- BIOSINTESIS PROTlanjutanDocument31 pagesBIOSINTESIS PROTlanjutanAprilikkaearlyNo ratings yet
- Metabolism of Aminoacids 2Document68 pagesMetabolism of Aminoacids 2Mi PatelNo ratings yet
- C10 Protein and Amino Acid MetabolismDocument8 pagesC10 Protein and Amino Acid MetabolismSoraya D. Al-ObinayNo ratings yet
- Urea CycleDocument6 pagesUrea CycleMegan GohNo ratings yet
- Amino Acid Metabolism All LecturesDocument18 pagesAmino Acid Metabolism All Lecturesmizare29gNo ratings yet
- B. Katabolisme Asam Amino-1Document19 pagesB. Katabolisme Asam Amino-1M Sifal MaulanaNo ratings yet
- Amino Acid MetabolismDocument97 pagesAmino Acid MetabolismEmmaNo ratings yet
- Answer B: A Deficiency in Carbamoyl Phos-: Metabolic BiochemistryDocument25 pagesAnswer B: A Deficiency in Carbamoyl Phos-: Metabolic BiochemistryHoanNo ratings yet
- Lecture Notes Urea Cycle Biochemistry 2, 2016 FV: Non-Essential Amino Acids Nitrogen MetabolismDocument34 pagesLecture Notes Urea Cycle Biochemistry 2, 2016 FV: Non-Essential Amino Acids Nitrogen MetabolismChaitali Dongaonkar ChawareNo ratings yet
- Amino Acid Catabolism1Document5 pagesAmino Acid Catabolism1rolwinNo ratings yet
- Amino Acid Metabolism NotesDocument152 pagesAmino Acid Metabolism NotesAkhilesh TiwariNo ratings yet
- Amino Acid Metabolism 2Document97 pagesAmino Acid Metabolism 2EmmaNo ratings yet
- Inborn Errors of Urea SynthesisDocument14 pagesInborn Errors of Urea SynthesisDaryl Jacob Bigay100% (1)
- Amino Acid MetabolismDocument10 pagesAmino Acid MetabolismatifzeaNo ratings yet
- Protein 3Document39 pagesProtein 3امجد حسين جواد كاظمNo ratings yet
- BP U9d Protein MetabolismDocument80 pagesBP U9d Protein MetabolismChristian Angelo AgbunagNo ratings yet
- Metabolism of Essential AADocument7 pagesMetabolism of Essential AA48 Syeda KainatNo ratings yet
- Bioquimica - Oxidación de Aminoácidos y Producción de UreaDocument51 pagesBioquimica - Oxidación de Aminoácidos y Producción de UreaLidia Escutia GuadarramaNo ratings yet
- Untitled 3Document38 pagesUntitled 3Reforwandis inagaNo ratings yet
- 3rd Module 1Document44 pages3rd Module 1KARTHIK VINODNo ratings yet
- Biochemistry Homework HelpDocument10 pagesBiochemistry Homework HelpassignmentpediaNo ratings yet
- Lectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-201-231Document31 pagesLectures - Biochemistry 1 - 2021-2022 Prof Version 6.01 - 23-24-201-231Ken M'voulaboloNo ratings yet
- Introduction Amino Acid Matabolism and CatabolismDocument45 pagesIntroduction Amino Acid Matabolism and CatabolismAboubakar Moalim Mahad moh'd100% (1)
- B314 AssignmentDocument23 pagesB314 AssignmentPalesa NtsekalleNo ratings yet
- Protein MetaboilisamDocument18 pagesProtein MetaboilisamSumit PandyaNo ratings yet
- AA Metabolism HANDOUTDocument4 pagesAA Metabolism HANDOUTnajwa hajjaliNo ratings yet
- Amino Acid Catabolism-Part-1: Biochemistry For Medics - Lecture Notes Professor (DR.) Namrata ChhabraDocument43 pagesAmino Acid Catabolism-Part-1: Biochemistry For Medics - Lecture Notes Professor (DR.) Namrata Chhabrashree devNo ratings yet
- General Protein Metabolism NUB Full 2022Document11 pagesGeneral Protein Metabolism NUB Full 2022PIH SHTNo ratings yet
- Urea CycleDocument39 pagesUrea Cycledrismailkm20No ratings yet
- Amino Acid MetabolismDocument25 pagesAmino Acid MetabolismParixit BhandurgeNo ratings yet
- Answers of Exam Ques.Document3 pagesAnswers of Exam Ques.Sonali SehrawatNo ratings yet
- Amino Acid Catabolism3Document2 pagesAmino Acid Catabolism3rolwinNo ratings yet
- Protein MetabolismDocument24 pagesProtein Metabolismstarboigaming08No ratings yet
- Presented by "Prof. Dr. Masoom Khan" Lecture Notes. No. 678Document41 pagesPresented by "Prof. Dr. Masoom Khan" Lecture Notes. No. 678Qamar QamarNo ratings yet
- Biochemistry Unit 03 PDFDocument66 pagesBiochemistry Unit 03 PDFDhanush kannanNo ratings yet
- BCH103 7Document6 pagesBCH103 7PRATEEK KUMARNo ratings yet
- Biochemistry MCQsDocument21 pagesBiochemistry MCQsnikteshgNo ratings yet
- All 20 of The Amino Acids Present in Proteins Are Essential For HealthDocument9 pagesAll 20 of The Amino Acids Present in Proteins Are Essential For HealthDeepu ChaurasiyaNo ratings yet
- General Reaction of Amino Acid BreakdownDocument20 pagesGeneral Reaction of Amino Acid BreakdownjessicatieuuNo ratings yet
- 03 - Metabolisme Protein - MonogastrikDocument25 pages03 - Metabolisme Protein - MonogastrikHerni Bustam100% (1)
- PROTEIN METABOLISM Dea Farha Fira Darson FineDocument44 pagesPROTEIN METABOLISM Dea Farha Fira Darson FineFarhati MardhiyahNo ratings yet
- Katabolisme Protein Dan Asam AmimoDocument35 pagesKatabolisme Protein Dan Asam AmimoLha YhoeLaNo ratings yet
- Protein MetabolismDocument19 pagesProtein MetabolismAnn Michelle Tarrobago100% (1)
- Urea CycleDocument3 pagesUrea CycleSundaralingam RajNo ratings yet
- Non-Essential Amino Acids Nitrogen MetabolismDocument21 pagesNon-Essential Amino Acids Nitrogen Metabolismpradeep36No ratings yet
- Amino Acid MetabolismDocument15 pagesAmino Acid Metabolismshanto.tn98No ratings yet
- BCH103 8Document6 pagesBCH103 8kingf3451No ratings yet
- Steam Ibr PDFDocument1 pageSteam Ibr PDFrolwinNo ratings yet
- AVEVA E3D Has In-Canvas CommandsDocument4 pagesAVEVA E3D Has In-Canvas CommandsrolwinNo ratings yet
- Utility Stations: Piping GuideDocument1 pageUtility Stations: Piping GuiderolwinNo ratings yet
- Piperack: PIPING GUIDE 02/04/03Document1 pagePiperack: PIPING GUIDE 02/04/03rolwinNo ratings yet
- STEAM Insl PDFDocument1 pageSTEAM Insl PDFrolwinNo ratings yet
- Basic Distillation Equipment and OperationDocument2 pagesBasic Distillation Equipment and OperationrolwinNo ratings yet
- New Kitchen by Karen MartiniDocument273 pagesNew Kitchen by Karen Martinirolwin100% (2)
- What'S New in Smartplant 3D: Product SheetDocument2 pagesWhat'S New in Smartplant 3D: Product SheetRaghu VenkatNo ratings yet
- Microsoft Windows Shortcut Keys List: AdvertisementDocument5 pagesMicrosoft Windows Shortcut Keys List: AdvertisementrolwinNo ratings yet
- Jadu File - Excel HelpDocument207 pagesJadu File - Excel HelpfecitusNo ratings yet
- SmartPlant3D 000Document2 pagesSmartPlant3D 000Ahmet_Cullu_681No ratings yet
- Amino Acid BiosynthesisDocument1 pageAmino Acid BiosynthesisrolwinNo ratings yet
- Design, Layout&drawing PDFDocument17 pagesDesign, Layout&drawing PDFrolwinNo ratings yet
- Astronomy - May 2018 USADocument80 pagesAstronomy - May 2018 USAJesús Gerardo Rodríguez FloresNo ratings yet
- Sp3d InstallDocument232 pagesSp3d Installrolwin50% (2)
- Keyboad 2 Modified 1 1Document7 pagesKeyboad 2 Modified 1 1leslie010No ratings yet
- A Guide To Refinery ProcessDocument35 pagesA Guide To Refinery ProcessBalamurugan100% (3)
- Releaase BuletineDocument56 pagesReleaase BuletinerolwinNo ratings yet
- SP3DInstall Checklist PDFDocument7 pagesSP3DInstall Checklist PDFrolwinNo ratings yet
- Flow Measurement Design Criteria - Oct 2015Document18 pagesFlow Measurement Design Criteria - Oct 2015rolwinNo ratings yet
- Amino Acid Catabolism3Document2 pagesAmino Acid Catabolism3rolwinNo ratings yet
- Amino Acid Catabolism1Document5 pagesAmino Acid Catabolism1rolwinNo ratings yet
- SM3DUpgradeGuide PDFDocument49 pagesSM3DUpgradeGuide PDFrolwinNo ratings yet
- Orifice Plate Fabrication Details - April 2014Document8 pagesOrifice Plate Fabrication Details - April 2014rolwinNo ratings yet
- SmartPlant InstructionsDocument5 pagesSmartPlant InstructionsrolwinNo ratings yet
- EquipmentDocument28 pagesEquipmentsathya2644No ratings yet
- DistillColumn enDocument54 pagesDistillColumn enrolwinNo ratings yet
- Heat Pipe Heat ExchangerDocument20 pagesHeat Pipe Heat ExchangerrolwinNo ratings yet
- How To Design Piping For Reboiling SystemsDocument7 pagesHow To Design Piping For Reboiling SystemsSalim YilmazNo ratings yet
- Test Bank For Microbiology 2nd Edition by Wessner Chapter 8 Not IncludedDocument25 pagesTest Bank For Microbiology 2nd Edition by Wessner Chapter 8 Not Includeda195840229100% (3)
- Glycolysis and PPP and Their Role in MetabolismDocument40 pagesGlycolysis and PPP and Their Role in Metabolismfak ouoNo ratings yet
- PCDC SummaryDocument6 pagesPCDC SummaryIsaac MwangiNo ratings yet
- Carbohydrates and Related Compounds: Unit 3Document19 pagesCarbohydrates and Related Compounds: Unit 3Gerald AquinoNo ratings yet
- M. F. McCarty, J. C. Gustin - Pyruvate and Hydroxycitrate/carnitine May Synergize To Promote Reverse Electron Transport in Hepatocyte Mitochondria, Effectively Uncoupling' The Oxidation of Fatty AcidsDocument10 pagesM. F. McCarty, J. C. Gustin - Pyruvate and Hydroxycitrate/carnitine May Synergize To Promote Reverse Electron Transport in Hepatocyte Mitochondria, Effectively Uncoupling' The Oxidation of Fatty AcidsAntonio Di DioNo ratings yet
- Gluconeogenesis, Glycogen MetabolismDocument35 pagesGluconeogenesis, Glycogen MetabolismdanielachynaNo ratings yet
- Bacterial MetabolismDocument46 pagesBacterial MetabolismMakarios S. YousefNo ratings yet
- Glycolysis Notes B.pharm 2ND Sem PDFDocument4 pagesGlycolysis Notes B.pharm 2ND Sem PDFBharti sain100% (2)
- Article OkDocument8 pagesArticle OkLK RismaNo ratings yet
- VCAA 2016 - Biology Examiner's ReportDocument11 pagesVCAA 2016 - Biology Examiner's ReportBob VNo ratings yet
- Solutions CH 13Document27 pagesSolutions CH 13Prinka AriessaNo ratings yet
- Kyambogo Report 2Document9 pagesKyambogo Report 2Nkugwa Mark WilliamNo ratings yet
- ACS Biochemistry Study PrepDocument13 pagesACS Biochemistry Study PrepJOHN100% (2)
- Biochemistry Revision E6.5Document72 pagesBiochemistry Revision E6.5Riya SharmaNo ratings yet
- Bioenergetics HandoutDocument1 pageBioenergetics HandoutRhaven GonzalesNo ratings yet
- Pyruvate Into Lactate and Back - From The Warburg Effect To Symbiotic EnergyDocument5 pagesPyruvate Into Lactate and Back - From The Warburg Effect To Symbiotic EnergyMirestean CamilNo ratings yet
- Backup of Goljan TransciptsDocument178 pagesBackup of Goljan TransciptsSarah RyanNo ratings yet
- Life Science Módulo 2Document48 pagesLife Science Módulo 2smokiermuffin83No ratings yet
- Lactobacillus SPP.: Lactobacillus Plantarum: Taxonomy, Identification, and Genetics of The SpeciesDocument8 pagesLactobacillus SPP.: Lactobacillus Plantarum: Taxonomy, Identification, and Genetics of The SpeciesAlex CárdenasNo ratings yet
- General BiologyDocument82 pagesGeneral BiologyNanashiNo ratings yet
- Curs 12 GlycolysisDocument48 pagesCurs 12 GlycolysisTeodora MunteanuNo ratings yet
- Locally Fermented Foods in Nigeria and Their Significance To National EconomyDocument12 pagesLocally Fermented Foods in Nigeria and Their Significance To National EconomyMonica EmeraldaNo ratings yet
- Aerobic and Anaerobic RespirationDocument7 pagesAerobic and Anaerobic RespirationFatmata Haja KamaraNo ratings yet
- 12~19 생화학 테뱅 정리Document117 pages12~19 생화학 테뱅 정리조주상No ratings yet
- Niger ATCC9142Document9 pagesNiger ATCC9142LiliOrangeLópezNo ratings yet
- Phenol-Red Carbohydrate Fermentation Broths PDFDocument3 pagesPhenol-Red Carbohydrate Fermentation Broths PDFLam Thanh NguyenNo ratings yet
- Christian Andersson Thesis-Succinic PDFDocument142 pagesChristian Andersson Thesis-Succinic PDFprivaz81No ratings yet
- Lactic Acid BacteriaDocument280 pagesLactic Acid BacteriaNicolas CalderonNo ratings yet
- Biochemistry Intermediate Exam Short Answers & CasesDocument14 pagesBiochemistry Intermediate Exam Short Answers & CasesMohammed EljackNo ratings yet